Variation in energy reserve in Rana tigrina and Bufo melanostictus during annual seasonal cycle
Amjad Fatmi1 , S. Nazre Ahsan2 and Durre Shahwar Ruby3
1
Department of Zoology,
Government P.G. College,
Dholpur,
India
2
Department of Zoology,
Patna University,
Patna,
India
3
Department of Zoology,
B.S. College Danpur,
Patna,
India
http://dx.doi.org/10.12944/CWE.4.1.31
Blood glucose, liver and muscle glycogen total serum protein and fat body are estimated during four phases of their annual. Liver glycogen was high during period of dormancy but low during breeding. Fat body weight was high during hibernation but low during post breeding season. Serum protein was high during hibernation and aestivation but low during breeding and post breeding seasons
Copy the following to cite this article:
Fatmi A, Ahsan S.N, Ruby D.S. Variation in energy reserve in Rana tigrina and Bufo melanostictus during annual seasonal cycle. Curr World Environ 2009;4(1):187-190 DOI:http://dx.doi.org/10.12944/CWE.4.1.31
Copy the following to cite this URL:
Fatmi A, Ahsan S.N, Ruby D.S. Variation in energy reserve in Rana tigrina and Bufo melanostictus during annual seasonal cycle. Curr World Environ 2009;4(1):187-190. Available from: http://www.cwejournal.org/?p=935
Download article (pdf)
Citation Manager
Publish History
Introduction
Amphibians are ectothermal vertebrates and like other poikilotherms are markedly influenced by seasonal change in temperature and water availability. Rana Tigrina and Bufo Melanostictus are two common Indian anurans. They under go four different phases during their annual biological cycle. Hibernation or winter sleep (Dec.-Feb.) and aestivation or summer torpidity (April-June) are periods of dormancy. During these periods they maintain minimum metabolic activities, practically starve and rely on energy reserves. The spawning seasons coincides with the monsoon rain from June to September (Mukherjee and Deb, 1960). Post breeding (Oct.-Nov.) is a hyperphagic phase during which large amount of food is consumed.
Glycogen in muscle and liver, fat in the fat bodies and serum proteins are major energy reserves in these vertebrates. There are some other less important energy reserves that could not be taken into account. Very few investigators have studied the metabolism of these energy reserves in anurans (Chaudhary 1984, Scapin et al., 1994).
In the present investigation, liver and muscle glycogen, Fat and fat bodies, serum protein and blood glucose has been investigated in both Rana tigrina and toad Bufo melanostictus in order to explore the role of these energy reserves in their physiological activities during the annual cycle.
Material and Methods
Normal healthy adult frog Rana tigrina and toad Bufo melanostictus were captured from Khagaul near Patna. Each toad wighed an average 150 gm. and a mean tip of snout to vent measurement of 90 mm. Each frog weighed an average of about 300 gm. and had a mean snout to vent measurement of 120 mm.
Collection where made in all the four season and each experiment sample consists of ten animals of each were taken for investigation for each seasons or phase. Quantitative estimation of total serum protein was made by the method of Kingsley (1942) followed by Mehl (1945) and Weichselbaum (1948). Blood glucose was estimated quantitatively by O-toluidine method of Cooper and Mc Danile (1970).
The quantitative estimation of glycogen from liver and muscle was done according to a modified method of kemp and Andrien (1954). Fat body was weighed by usual method.
Results and Discussion
In both Rana tigrina and Bufo melanostictus, blood glucose level in low during hibernation and aestivation, but liver glycogen is higher during these period, low level of blood glucose may be related to its relatively inactive nature of these anurans during these periods and the surplus of glucose is converted into glycogen in the liver and serves as energy reserves. Wintering in ectothermal vertebrates causes low blood glucose and high lives glycogen (Bonnet 1994, Scapin et al., 1994 Silvia et al., 2004). The high liver glycogen found in the present study may also be due to the fact that during these period both the animals practically starve. The high glycogen content of the liver is associated with significant drop in feeding. Liver glycogen content is grater during hibernation than summer. Freeze Tolerence in various amphibians promote production of glucose in the form of cryoprotectant by glycogenolysis in hepatocytes (Holden et al., 2000, Mommsen et al., 1992). The view supports utilization of liver glycogen during the period between hibernation and aestivation.
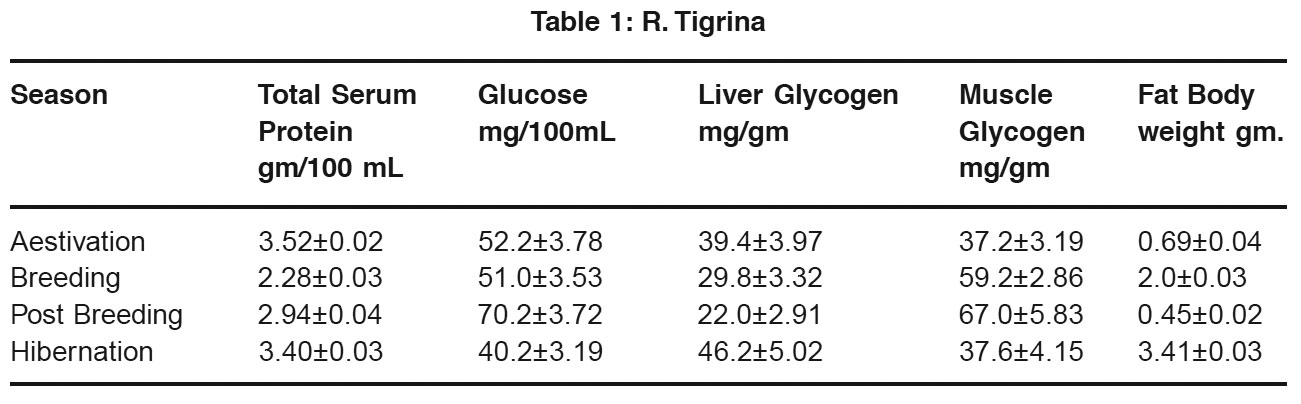 |
Table 1: R. Tigrina Click here to view table |
In both Rana and Bufo blood glucose and liver glycogen content is low during breeding season. It is due to increased utilization of this energy reserve in spawning for energy need. A higher level of liver glycogen content inBufo than in Rana is indicative of a grater dependence of the former on liver glycogen than the later.
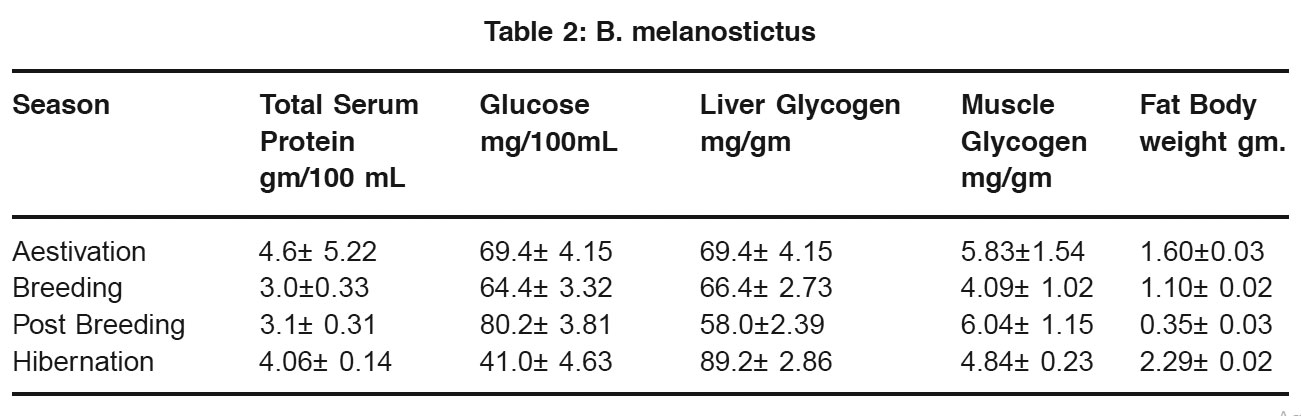 |
Table 2: B. melanostictus Click here to view table |
In both the anurans studied there is a slight increase muscle glycogen during summer as compared hibernation. The muscle glycogen store probably increased at the cost of liver glycogen because liver glycogen is lower and blood glucose is higher during summer than winter. The increase in muscle glycogen during this period may be seem as an adaptation towards increased nee in locomotor activity during the following seasons i.e. spawning season. Muscles glycogen decreases during hibernation although liver glycogen and fat bodies are high. It is possible that muscle glycogen rather than liver glycogenm is the major source of blood glucose during hibernation and also because locomotor activity considerably dropped muscle glycogen is transported from muscle to liver and get stored in liver.
In poikilotherms, fat bodies are the main depot for lipid (Dickerson 1976, Lela et al., 1979). In both Rana and Bufo fat body weight is highest during hibernation. It is due to dormancy and low BMR (Flanengan 1993, Tocque 1994). In both the amphibians fat body weight is very low immediately after breeding. There is a rapid rise in the size and weight of the fat body from post breeding to hibernation. The rapid increase in fat body weight is obviously related to intensity of feeding during post breeding season. In Rana there is a marked decrease in fat body weight. In the period between hibernation and aestivation but the slope in Bufo is less steeper. Although metabolism in hibernating amphibians are low but synthetic activities continue. This reflects the difference in extent of individual reserves found during hibernation and aestivation. In tegu lizard during dormancy fatty acids from fat is converted into β – hydroxyl butyric acid in liver to compensate for glucose as source of energy (Stuart and Vallentyne 1997). This view also support utilization of fat during these periods by poikilitherms. In both Rana and Bufo serum protein level is higher during aestivation and hibernation and there is light increase from hibernation to aestivation. This indicates that during period of torpidity, these two amphibians utilize mainly lipid and glycogen as energy reserves and maintain a high level of protein. Epigian and hypogian amphibian show variation in dergree of utilization of individual energy reserves (Harvent et al., 2001). From the preset study it is revealed that in Rana there is not much difference in liver glycogen during hibernation and aestivation but fat body weight is much lower in summer than winter. In Bufo liver glycogen content is much less in aestivation than hibernation and there is not much difference in weight of fat bodies. This indicates that Rana primarily utilizes fat store for obtaining energy needed during winter and summer and most of the glucose in blood is of gluconeogenic origin where as Befo mostly utilizes glycogen as source of blood glucose during these periods. In Rana there is accretion of fat bodies from aestivation to breeding but in Bufo, there is a slight decrease in fat bodies during this period. Glycogen content and serum protein decreases from aestivation to breeding but the slope of glycogen level is more steeper in frog than in toad. This indicates that Rana primarily utilizes glycogen and protein for immediate energy needs and accumulates fat for their utilization during breeding season. Whereas Bufo utilizes protein, glycogen and fat for energy requirement during this period.
The fat body weight is minimum in both Rana and Bufo during post breeding period. It is related to the fact that the fat bodies are utilized primarily for reproductive activities, gametogenesis etc. during breeding season (Selvia et al., 2004). The range of variation in fat body weight is markedly higher in Rana than in Bufo which probably suggests a grater dependence on fat for energy by Rana than Bufo.
References
1. Bonnet X., naulleau, G., Relationship between glycemia and seasonal activity in Vipera aspis, L. Amphibia Reptilia. (1994) 14: 295-306.
2. Cooper, G.R., V. Mcdanile, Manual of routine methods in Clinical Chemistry for use in intermediate laboratories. Std. mehods Clin.Chem. (1970) 6: 159-170.
3. Kemp. A.: J. Andreien., A colorimetric micro method for the determination of glycogen intissue, Biochem. J. (1954) 56: 646-648.
4. Kingsley, A colorimetric method of blood analysis, In Hawk’s physiological chemistry. Oser. B. III (ed) Tata McGraw Hill. New Delhi (1942).
5. Harvent if: Mathieu J. Durand J., Behavioral, physiological and metabolic responses to long term starvation and re-feeding in a blind cave dwwlling (proteus Anguinus) and a surface dwelling (Euproctus asper) salamander, J. Exp. Biol. Jan; (2001) 204(Pt2): 269-81.
6. Holden, C.P., Storey, K.B., Purification and characterization of protein Kinase A from liver of the freeze tolerant wood frog: role in glycogenolysis during freezing. Cryobiology. Jun: (2000) 40(4): 323-31.
7. Mommsen, TP; Story KB., Hormonal effect on glycogen metabolism in isolated hepatocytes of a freeze- tolerant frog. Gen. Comp. Endocrinol. Jul; (1992) 8791: 44-53.
8. Scapin S.; Di Giuseppe, G., Seasonal variation of glycogen synthetase and phosphorylase activities in the frog Rana Esculenta. Comp. Biochem. Physiol. B (1994) 107:189-195 .
9. Stuart, J.A.; Ballantyne, J.S., Importance of Ketone bodies to the intermediary metabolism of the terrestrial snail, Archachartina Ventricosa; abidance from enzyme activities. Comp. Biochem. Physiol. B. (1997) 117: 197-201.
10. Silvia Cristina; JED Carvalho; AS. A. BE; J.E.P.W. Bicudo. M.S.C. Bianconcini., Seasonal metabolic depression, substrate utilization and changes in scaling patterns during the first year cycle of tegu lizard (Tupinambis merianae). J. Exp. Biol. (2004) 207: 307-318.







