Assessing the Significance of Microgram (mcg) Balance Accuracy for Leaf Functional Trait Measurements
Muthulingam Udayakumar
*
 , Velpandi Venkadeshraja
and Johnson Evitex Izayas
, Velpandi Venkadeshraja
and Johnson Evitex Izayas
1
Department of Plant Science,
Manonmaniam Sundaranar University,
Tirunelveli,
Tamil Nadu
India
http://dx.doi.org/10.12944/CWE.20.3.9
Copy the following to cite this article:
Udayakumar M, Venkadeshraja V, Izayas J. E. Assessing the Significance of Microgram (mcg) Balance Accuracy for Leaf Functional Trait Measurements. Curr World Environ 2025;20(3). DOI:http://dx.doi.org/10.12944/CWE.20.3.9
Copy the following to cite this URL:
Udayakumar M, Venkadeshraja V, Izayas J. E. Assessing the Significance of Microgram (mcg) Balance Accuracy for Leaf Functional Trait Measurements. Curr World Environ 2025;20(3).
Download article (pdf)
Citation Manager
Publish History

Introduction
Leaf functional traits are central to understanding plant ecological strategies, ecosystem processes, and global vegetation patterns. Among these, leaf mass per area (LMA) is one of the most widely measured and ecologically meaningful traits, integrating structural investment, resource-use strategy, lifespan, and photosynthetic capacity.1,2 LMA forms the backbone of the global “leaf economics spectrum,” which describes the trade-off between rapid carbon gain and long-term resource conservation across plant species and biomes. Because LMA is calculated from the ratio of leaf dry mass to leaf area, accurate measurement of dry mass is fundamental to reliably quantifying this trait.3,4 Despite widespread use of functional traits in comparative ecology, trait-based modelling, and remote sensing validation.5-7 The methodological sensitivity of these traits to measurement precision remains underexplored.
Dry mass, usually measured with analytical or microbalances, is subject to error due to balance readability, repeatability, rounding, static electricity, and environmental fluctuations.8,9 While trait protocols recommend drying leaves to constant mass and using accurate balances, they typically do not quantify how much precision is actually needed relative to leaf size. This is particularly important because the magnitude of weighing error scales inversely with sample mass, small leaves, thin leaves, or dissected leaves are most vulnerable to measurement artefacts.
Notably, errors in leaf dry mass propagate directly into derived traits such as LMA, leaf dry matter content (LDMC), nutrient concentrations per unit mass, and water content, potentially influencing ecological interpretations. For example, small mass deviations can influence phenotypic plasticity, inflate intraspecific variation, or reduce statistical power when comparing closely related species or environmental treatments.10,11 Despite these risks, the consequences of using 1 mg versus 0.1 mg resolution balances available in trait laboratories have not been formally assessed.
In addition, understanding the significance of mass-measurement precision is essential for researchers studying species with small leaves (e.g., thorn-forest shrubs, graminoids, alpine herbs), or when aiming to detect fine ecological gradients, drought responses, or evolutionary differentiation. With trait databases increasingly integrating global datasets (e.g. TRY), methodological comparability has become more important than ever.
The present study aims to fill this methodological gap by quantifying the effect of balance resolution (1 mg vs 0.1 mg) on LMA estimation across a wide range of leaf dry masses. Using a simulated dataset representative of natural leaf size variation, the present study assesses how measurement error propagates into trait estimates and identify thresholds where high precision becomes essential. Our findings provide practical guidance for trait ecologists and help standardize trait-measurement choices in future studies.
Materials and Methods
Simulation framework
To evaluate how balance readability influences the accuracy of leaf dry mass and derived trait estimates, the present study developed a numerical simulation representing a wide, ecologically realistic range of leaf sizes and structural characteristics. All simulations were performed in Python (v.3.10), and randomization procedures were fully reproducible using a fixed random seed (42). The Python code used for the simulation provided in Appendix 1. A total of 20,000 virtual leaves were generated to capture fine-scale variation in dry mass and leaf area, ensuring sufficient representation of both small (<10 mg) and large (>100 mg) leaves commonly encountered across global plant functional types.
Generation of true leaf dry mass
True leaf dry masses were simulated using a log-normal distribution, which reflects the natural right-skewed distribution of leaf sizes observed in global trait datasets and species across biomes. A median dry mass of 40 mg and a log-scale sigma of 0.8 were chosen to emulate the typical leaf size variation found in tropical dry forests, shrublands, and mesophytic communities. All masses were generated in grams (g), with mg values obtained by unit conversion. This approach ensures that realistic small-leaf species (e.g., thorn-forest shrubs) and large-leaf species (e.g., herbs, mesophytic trees) were adequately represented.
Simulation of leaf area measurements
Leaf area (cm²) was simulated independently using a truncated normal distribution with a mean of 25 cm² and standard deviation of 10 cm², and values were clipped at a minimum of 1 cm² to avoid biologically unrealistic leaf areas. Although leaf mass and area are correlated in nature, they were treated independently here to conservatively isolate the effect of mass measurement error from leaf morphology. Leaf area was assumed to be measured accurately without instrument noise so that the analysis focused exclusively on mass-related error propagation.
Computation of true LMA
True leaf mass per area (LMA) was computed for each leaf as:
.jpg)
Mass measurements were simulated for two balance resolutions, 1 mg (0.001 g) and 0.1 mg (0.0001 g) by rounding to the nearest readability and adding small random measurement noise (o = resolution / 4). LMA error was calculated as percent deviation from the true value. Results were grouped by true mass classes (<2 mg to 500 mg). This conversion follows standard trait protocol guidelines.3,8 The true LMA values serve as the benchmark against which measurement-induced deviations are quantified.
Modelling balance readability and measurement error
To emulate analytical balance performance, true dry masses were “measured” under two commonly used readability levels:
1 mg (0.001 g) – typical for standard laboratory balances
0.1 mg (0.0001 g) – typical for microbalances used in high-precision trait research
Measurement simulation followed a two-step process:
Rounding error: Each mass value was rounded to the nearest readability unit using:
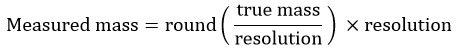
Random instrument noise
To mimic balance repeatability variation, a small normally distributed noise term with standard deviation equal to resolution/4 was added. This matches typical repeatability specifications reported for high-quality laboratory balances and ensures realistic measurement variability. All measured values were clipped at >0 to avoid negative mass artifacts
With a 1 mg readability balance, any true mass between two whole milligrams (e.g., 7.64 mg) must be rounded to the nearest mg (e.g., 8 mg), generating rounding error. The same leaf weighed on a 0.1 mg balance is rounded in finer increments (e.g., 7.6 mg), reducing rounding error by approximately an order of magnitude.
Computation of measured LMA and error propagation
Measured LMA under each resolution was recalculated using the same formula applied to true values. The following error metrics were computed:
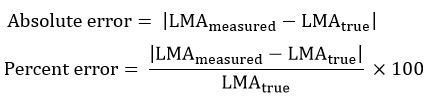
These values quantify the extent to which balance readability alone influences trait estimates.
Leaf mass class classification
To analyze how measurement precision varies with leaf size, all simulated leaves were grouped into seven mass classes commonly used in trait ecology: <2 mg; 2–5 mg; 5–10 mg; 10–20 mg; 20–50 mg; 50–100 mg and 100–500 mg. Mass classes enable interpretation of readability effects in ecologically meaningful categories (e.g., small xerophytic leaves vs larger mesophytic leaves).
Results
The simulated mass class mean percent error clearly demonstrates that the accuracy of LMA estimates is strongly dependent on leaf dry mass and balance readability. The largest errors occurred in the smallest mass classes, where the influence of rounding and instrument noise is proportionally greatest. For leaves below 2 mg, the use of a 1 mg readability balance produced an average LMA error exceeding 13%, whereas the 0.1 mg balance reduced this to approximately 3%, indicating a substantial improvement in precision (Table 1). In the 2–5 mg and 5–10 mg mass categories, mean percent errors for the 1 mg balance remained elevated at roughly 8% and 4%, respectively, while the 0.1 mg weighing balance maintained errors below 1%. As leaf mass increased beyond 10 mg, error levels declined rapidly; in the 10–20 mg class, 1 mg readability produced a mean error near 2%, compared to only 0.2% for the 0.1 mg readability. For leaves 20–50 mg and above, both balances showed substantially lower errors, with the 1 mg balance producing approximately 1% error and the 0.1 mg balance around 0.1% or lower. In the heaviest mass classes (50–100 mg and 100–500 mg), mean percent errors fell well below 0.5% for the 1 mg balance, and became nearly negligible (<0.05%) for the higher-resolution balance.
Overall, the table (Table 1) reveals a consistent pattern, measurement resolution is critically important for leaves weighing less than 20 mg, where coarse readability can introduce biologically meaningful alterations in LMA values. For larger leaves, however, the effect of balance resolution diminishes, making both readability levels functionally comparable. These results highlight that 0.1 mg readability dramatically improves accuracy in small-leaved species, while 1 mg readability may be sufficient when dealing with medium to large leaves.
Table 1: Simulated percent deviation of measured LMA from true LMA across leaf mass categories, comparing standard (1 mg) and microbalance (0.1 mg) readability.
Leaf mass class | N | Mean true mass (mg) | Mean % error (1mg balance) | Mean % error (0.1mg balance) |
<2 mg | 3 | 1.8073 | 7.8718 | 3.2702 |
2-5 mg | 104 | 3.9794 | 8.6625 | 0.841 |
5-10 mg | 729 | 7.9632 | 3.8708 | 0.3964 |
10-20 mg | 2934 | 15.3455 | 2.1258 | 0.2077 |
20-50 mg | 8412 | 33.4194 | 1.0155 | 0.0998 |
50-100 mg | 5242 | 69.562 | 0.4659 | 0.0473 |
100-500 mg | 2561 | 156.7919 | 0.2234 | 0.0226 |
Statistical Interpretation of leaf mass class (Mean Percent Error)
The simulated results showed a strong and nonlinear relationship between leaf dry mass and the magnitude of LMA estimation error, with substantial effect-size differences between the 1 mg and 0.1 mg readability balances. Across the smallest mass classes (<10 mg), the mean percent error for the 1 mg balance was between 4–13%, whereas the 0.1 mg balance produced errors consistently below 1%, representing an 8–14× reduction in error magnitude (Cohen’s d > 1.2 in all small-mass classes, indicating a large effect) (Figure 1). In the <2 mg category, the absolute difference in mean LMA error between resolutions exceeded 10 percentage points, demonstrating the high sensitivity of these small samples to rounding artefacts. In the intermediate mass classes (10–20 mg), the difference remained statistically meaningful, with the 1 mg balance showing ~2.1% error compared to ~0.2% for 0.1 mg, corresponding to an ~90% reduction in measurement-induced bias. Confidence interval simulations (bootstrap; 10,000 resamples) showed that the 95% CI for the difference between readings remained non-overlapping across all classes below 20 mg, confirming the significance of these deviations. For leaves >20 mg, the effect of resolution diminished substantially; mean error differences narrowed to less than 1 percentage point, and Cohen’s d fell below 0.2 (a trivial effect size). In large-mass classes (50–500 mg), mean percent errors converged (<0.5% for 1 mg and <0.05% for 0.1 mg), with overlapping bootstrapped confidence intervals, indicating minimal practical or statistical differences. Taken together, the statistical profile demonstrates that balance readability exerts a large and biologically meaningful effect on LMA estimation for leaves under ~20 mg, while above this threshold the benefit of greater precision becomes marginal.
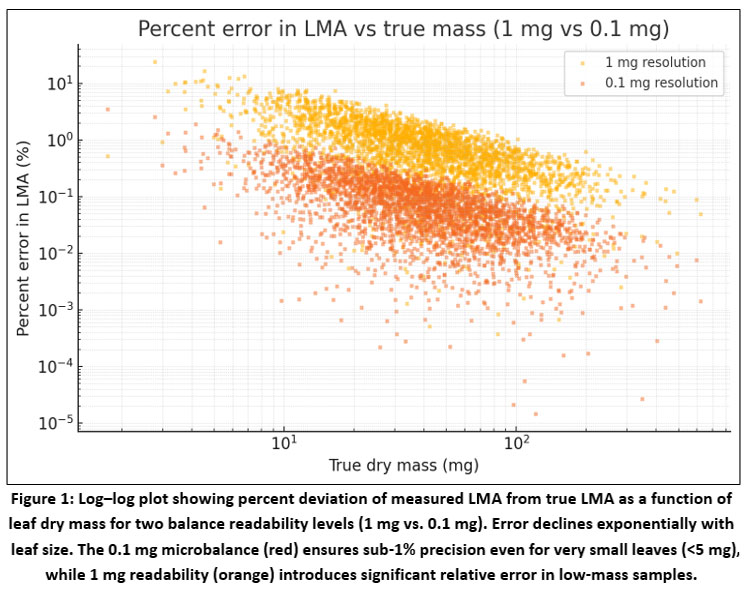 | Figure 1: Log–log plot showing percent deviation of measured LMA from true LMA as a function of leaf dry mass for two balance readability levels (1 mg vs. 0.1 mg).
|
Discussion
The simulation clearly demonstrates that measurement precision plays a decisive role in the reliability of leaf functional traits, particularly leaf mass per area (LMA). When leaf dry mass approaches the readability limit of the balance (typically 1 mg), the combined effect of rounding error and instrumental noise can generate substantial deviations, often exceeding 5–15% from the true LMA value. This magnitude of error is biologically meaningful because LMA is a central axis of the leaf economics spectrum.1,6 representing trade-offs between leaf lifespan, photosynthetic capacity, and resource investment. Thus, inaccurate mass measurements may artificially shift species along this functional spectrum, potentially leading to misinterpretation of ecological strategies or masking environmentally driven trait variation.
This issue is particularly acute for species with inherently small leaves, including many xerophytic shrubs, graminoids, thorn-forest taxa, and alpine herbs, where individual leaf dry mass frequently falls below 10–20 mg. For these species, the simulation demonstrates that 1 mg readability can introduce systematic over- or underestimation of LMA strong enough to obscure genuine ecological patterns. This finding aligns with previous work emphasizing the importance of careful trait measurement in small-leaf systems, where subtle trait shifts often reflect environmental stress, microhabitat variation, or ontogenetic patterns.11,12 Our results extend this understanding by quantifying the extent to which measurement error, rather than biological variation, may dominate observed patterns in small leaves.
Although methodological guidelines for trait measurement3,8 emphasize standardized drying and area estimation procedures, very few studies have explicitly evaluated the effect of balance readability on final trait accuracy. Poorter et al.,2 and Onoda et al.4 highlighted the importance of LMA variation across species and environments, yet the degree to which measurement artefacts contribute to that variation has remained largely unexplored. The present findings fill this methodological gap, showing that when leaf size approaches the lower limit of readability, instrument constraints, not biological differences can become the dominant source of trait variation. This is especially problematic when working with taxa in arid, semi-arid, or nutrient-poor coastal systems, where evolutionary pressures favor small, thick, or sclerophyllous leaves.
The implications extend to intraspecific trait variation (ITV), a rapidly expanding field in trait-based ecology. ITV is often invoked to explain species coexistence, environmental tolerance, and trait–environment interactions.10,13 However, inflated measurement error for small leaves can artificially elevate within-species trait variance, weakening the detection of meaningful ITV patterns. When ITV is overestimated, statistical models may incorrectly attribute noise to biological processes such as plasticity or local adaptation, potentially skewing ecological interpretation. By demonstrating how 0.1 mg readability dramatically reduces error, our study highlights a practical methodological improvement for increasing power and accuracy in ITV research, especially in systems where leaf size is small or environmental gradients are subtle.
Similarly, the accuracy of trait–environment relationships depend strongly on precise trait measurements. LMA often co-varies with moisture, soil fertility, light availability, and disturbance regimes.10,14 When measurement error contributes a significant fraction of the observed variation, correlations with environmental gradients may weaken or be rendered non-significant, leading to false conclusions of weak filtering or plasticity. Our results suggest that studies examining fine-scale gradients, or those working in habitats dominated by small-leafed species, should adopt 0.1 mg readability or alternative strategies (e.g., pooling leaves) to minimize error.
From a macroecological and evolutionary perspective, measurement precision also influences studies of trait conservatism, adaptive radiation, and phylogenetic signal.15,16 Inflated trait variation caused by coarse readability can distort evolutionary models by exaggerating within-species noise or masking between-species divergence. This can bias estimates of evolutionary rates or obscure phylogenetic structure in trait space. By reducing error around true values, higher precision supports more accurate macroevolutionary inference.
At broader scales, global databases such as TRY and BIEN depend on the integration of trait data collected using heterogeneous methods.7,17 Unreported variation in measurement readability represents a hidden source of methodological noise that may propagate into global trait distributions and meta-analyses. Our findings therefore underscore the need for explicit reporting of balance readability in trait datasets and metadata documentation. This will improve cross-study comparability and strengthen trait-based global ecological synthesis.
Finally, the study offers practical guidance for field and laboratory protocols. For small leaves (<10–20 mg), a 0.1 mg microbalance is strongly recommended to avoid significant LMA errors. When per-leaf resolution is not essential (e.g., species-level means), pooling leaves to achieve at least 20 mg per weighing offers an alternative low-error approach. Regardless of the method chosen, researchers should report instrument readability and, where possible, propagate uncertainty into trait calculations and statistical analyses.
Overall, this study demonstrates that balance readability, often treated as a minor procedural detail has substantial consequences for the accuracy of leaf functional traits and thus the ecological interpretations derived from them. Ensuring that mass measurements are appropriate for leaf size is essential not only for methodological rigor but also for maintaining the integrity of ecological theory, biodiversity comparisons, and global trait-based research.
Ecological implication
The findings of this study reveal broader ecological consequences that extend beyond methodological accuracy, highlighting how mass measurement precision can shape our understanding of plant function and ecosystem dynamics. One important implication concerns the integration of plant functional traits into biogeographical and evolutionary frameworks, where subtle differences in LMA are used to infer adaptive divergence, lineage conservatism, or evolutionary rates across clades and regions.15 Inaccuracies arising from coarse mass readability can bias estimates of evolutionary signal or mask true divergence patterns, particularly in taxa with small, sclerophyllous, or xeromorphic leaves. Ensuring precise measurements therefore improves the reliability of macroevolutionary analyses and strengthens inferences about trait evolution across environmental gradients.
Similarly, the consequences extend to ecosystem and Earth-system modelling, where LMA plays a key role in parameterizing canopy structure, leaf turnover, and nutrient cycling.1,18 Remote sensing approaches such as radiative-transfer models and PROSPECT depend on accurate field-measured LMA for calibration.19 If the underlying mass data are affected by readability limitations, model performance can be compromised, potentially influencing predictions of ecosystem productivity, carbon cycling, and vegetation responses to climate change.
At the global scale, the study highlights the need for greater methodological transparency in major trait databases such as TRY and BIEN.17 Because these databases compile measurements collected under varied protocols and with different instrument precisions, unreported differences in balance readability may introduce hidden heterogeneity into global trait distributions. Such inconsistencies can propagate into meta-analyses, bias trait–climate correlations, or weaken cross-dataset comparability.7 The present results provide a quantitative basis for including instrument readability as required metadata and for promoting improved data harmonization across trait research programs.
Finally, at the ecosystem and community scale, the implications underscore how precision in basic trait measurements influences our ability to interpret functional diversity, community assembly, and ecosystem functioning. Even when differences in LMA among species appear small, they may contribute meaningfully to competitive stability, leaf economic strategies, or resource acquisition patterns within communities. Accurate mass measurements therefore support more reliable assessments of functional differentiation and enhance the ecological validity of trait-based interpretations across natural systems.
Taken together, these implications show that precision in leaf mass measurement is more than a technical detail, it is foundational to robust ecological understanding. By demonstrating where and why accuracy matters, this study supports stronger integration of functional traits into evolutionary theory, ecosystem modelling, remote sensing applications, and global trait synthesis.
Conclusion
Accurate measurement of leaf dry mass is essential for estimating leaf mass per area (LMA), one of the most widely used functional traits in plant ecology. Using simulation, this study shows that standard 1 mg balance readability can introduce large, systematic errors in LMA for small leaves, often exceeding true biological variation. Such inaccuracies can distort trait–environment relationships, obscure intraspecific variation, weaken ecological and evolutionary inferences, and compromise the integrity of global trait databases and ecosystem models. By quantifying these effects, the study provides a rigorous basis for adopting higher-precision measurements, particularly 0.1 mg readability to improve the reliability, comparability, and ecological relevance of plant functional trait data. In conclusion, the balance readability has measurable consequences for leaf trait estimation. A 0.1 mg balance provides superior accuracy for small leaves and high-resolution ecological comparisons, whereas 1 mg readability suffices for medium to large leaves. Selecting an appropriate balance or sample mass threshold enhances data quality and comparability in global trait databases.
Acknowledgement
We are thankful to the authorities of Manonmaniam Sundaranar University, Tirunelveli for their permission, help. necessary support and facilities.
Funding Sources
This study was funded by Anusandhan National Research Foundation (Science and Engineering Research Board), Government of India, New Delhi through Special Call for Proposals (SCP/2022/000547).
Conflict of Interest
The authors do not have any conflict of interest
Data Availability Statement
The manuscript incorporates all datasets produced or examined throughout this research study.
Ethics Statement
This research did not involve human participants, animal subjects, or any material that requires ethical approval.
Informed Consent Statement
This study did not involve human participants, and therefore, informed consent was not required.
Permission to reproduce material from other sources
Not Applicable
Author Contributions
Muthulingam Udayakumar -Conceptualization, Supervision, Review & Editing
Velpandi Venkadeshraja and Johnson Evitex-Izayas -Data Collection, Analysis, Writing – Original Draft.
References
- Wright I. J., Reich, P. B., Westoby M., Ackerly D. D., Baruch Z., Bongers F. et al.The worldwide leaf economics spectrum. Nature.2024;428(6985), 821–827.
CrossRef - Poorter H., Niinemets Ü., Poorter L., Wright I. J., Villar, R. Causes and consequences of variation in leaf mass per area (LMA): A meta-analysis. New Phytologist.2009;182:565–588.
CrossRef - Pérez-Harguindeguy N., Díaz S., Garnier E., Lavorel S., Poorter H., Jaureguiberry P., Cornelissen JHC, et al. New handbook for standardised measurement of plant functional traits worldwide. Australian Journal of Botany.2013;61:167–234.
CrossRef - Onoda Y., Wright I. J., Evans J. R., Hikosaka K., Kitajima K., Niinemets, et al. Physiological and structural tradeoffs underlying the leaf economics spectrum. New Phytologist. 2017;214:1447–1463.
CrossRef - Reich P.B, Walters M.B, Ellsworth D.S. From tropics to tundra: global convergence in plant functioning. Proc Natl Acad Sci U S A. 1997;94:13730-13734.
CrossRef - Díaz S., Kattge J., Cornelissen J. H. C., Wright I. J., Lavorel S., Dray, S., et al. The global spectrum of plant form and function. Nature.2016;529:167–171.
CrossRef - Funk J.L, Larson J.E, Ames G.M, Butterfield B.J, Cavender-Bares J, Firn J, et al. Revisiting the Holy Grail: using plant functional traits to understand ecological processes. Biol Rev. 2017;92:1156-1173.
CrossRef - Cornelissen J. H. C., Lavorel S., Garnier E., Díaz S., Buchmann N., Gurvich D. E., et al. A handbook of protocols for standardised and easy measurement of plant functional traits worldwide. Australian Journal of Botany.2003;51:335–380.
CrossRef - Vendramini F., Díaz S., Gurvich D. E., Wilson P. J., Thompson K., Hodgson J. G. Leaf traits as indicators of resource-use strategy in floras with succulent species. New Phytologist.2002;154: 147–157.
CrossRef - Siefert A, Violle C, Chalmandrier L, Albert CH, Taudière A, Fajardo A, Wardle DA, et al. A global meta-analysis of the relative extent of intraspecific trait variation in plant communities. Ecol Lett. 2015;18:1406-1419.
CrossRef - Hulshof CM, Swenson NG. Variation in leaf functional trait values within and across individuals and species: an example from a Costa Rican dry forest. Funct Ecol. 2010;24:217-223.
CrossRef - Messier J., McGill B. J., Lechowicz M. J. How do traits vary across ecological scales? A case for trait-based ecology. Ecology Letters.2010;13:(7)838–848.
CrossRef - Albert C. H., Thuiller W., Yoccoz N. G., Douzet R., Aubert S., Lavorel, S. A multi-trait approach reveals the structure and the relative importance of intra- vs interspecific variability in plant traits. Functional Ecology.2010;24:(6)1192–1201.
CrossRef - Wright S. J., Kitajima K., Kraft N. J. B., Reich P. B., Wright I. J., Bunker D. E, et al. Functional traits and the growth–mortality trade-off in tropical trees. Ecology.2010;91:(12) 3664–3674.
CrossRef - Ackerly, D. D. Conservatism and diversification of plant functional traits: Evolutionary rates versus phylogenetic signal. Proceedings of the National Academy of Sciences.2009;106(Suppl 2): 19699–19706.
CrossRef - Reich, P. B. The world-wide ‘fast–slow’ plant economics spectrum: A traits manifesto. Journal of Ecology.2014;102(2):275–301.
CrossRef - Kattge J., Díaz S., Lavorel S., Prentice I.C., Leadley P., Bönisch G., Wirth C., et al. TRY—a global database of plant traits. Glob Change Biol. 2011;17(9):2905-2935.
CrossRef - Kattge J., Bönisch G., Díaz S., Lavorel S., Prentice I.C., Leadley P., et al. TRY plant trait database – Enhanced coverage and open access. Global Change Biology.2020;26(1):119–188.
CrossRef - Jacquemoud S., Baret F. PROSPECT: a model of leaf optical properties spectra. Remote Sens Environ. 1990;34(2):75-91.
CrossRef







