Regeneration Status of Quercus lanata (Syn. Q.lanuginosa) in Central Himalayan Forests
Geeta Sharma
*
 , Ashish Tewari
and Shruti Shah
, Ashish Tewari
and Shruti Shah
1
Department of Forestry,
DSB Campus, Kumaun University,
Nainital,
India
http://dx.doi.org/10.12944/CWE.20.2.10
Copy the following to cite this article:
Sharma G, Tewari A, Shah S. Regeneration Status of Quercus lanata (Syn. Q.lanuginosa) in Central Himalayan Forests. Curr World Environ 2025;20(2). DOI:http://dx.doi.org/10.12944/CWE.20.2.10
Copy the following to cite this URL:
Sharma G, Tewari A, Shah S. Regeneration Status of Quercus lanata (Syn. Q.lanuginosa) in Central Himalayan Forests. Curr World Environ 2025;20(2).
Download article (pdf)
Citation Manager
Publish History

Introduction
Oak forests are considered as a keystone species and inhabit an area around 20,000km in Central Himalaya.1 Himalaya is one of the most fragile ecosystems in the world because of its diverse, rough, rugged and undulating topographical features.2,3 In the Himalayan Forest, there have been numerous documented alterations in composition, density, structure, and regeneration patterns attributable to various biotic pressures. The area exhibits a range of climates, from subtropical to alpine, sustaining a broad range of plant and animal life. This climatic diversity, coupled with the varying elevations in the region, results in a unique and rich ecological environment.
Genus Quercus is native to northern hemisphere and encompasses deciduous and evergreen species, exhibiting range of latitudes from cool temperate to tropical. Majority of oak species are found on continents of Asia, Europe, America, and North Africa. Oaks in the Himalayan region significantly contribute to soil and water conservation, biodiversity maintenance, sustainability, and other ecosystem services.4 Oaks performs vital part in Indian Himalayas, serving as key sources of agricultural tools, fuelwood, fodder, charcoal, and materials for making beehive boxes. During dry or lean seasons, oak leaves are commonly used as livestock fodder and also as bedding material for animals.5 Additionally, oaks facilitate the replenishment of mountain springs.6
In Indian Himalayan region, moist temperate forests are dominated by oaks (Quercus spp.), which are climax tree species. Around 35 species of Quercus are widely distributed between 1000 to 3500m elevations.7 In Uttarakhand, there are five species of oaks, with the exception of Q. glauca and Q. lanata, all other oak species have widespread distribution and collectively constitute extensive regions in Himalaya.8 The anthropogenic pressure on oak forest are high as people depend on their daily needs of fuelwood and fodder on these oaks or the region.
Q. lanata (rianj) is a large evergreen tree and is usually low branched. It is an oak native to Northern Thailand, China, and Vietnam, where it grows at altitudes between 800 & 2600m In the hills of India, it is found at elevations ranging from 1800 to 2400m.9 It occurs in small patches in the Garhwal and Kumaun Himalaya. It has been associated with Rhododendron arboreum and Q. leucotrichophora at its lower limit and with Q. floribunda towards upper limit respectively.7 Very few studies on regeneration and seed ecology for Q. lanata.
The phytosociological information of a particular tree species helps to better understand their ecology, habit, habitat, vegetational structure, and how they are inter-connected.10,11 The basic goal of the forest ecologist is to understand the dynamics of landscape ecosystems, their structure and functions.12
The composition and dynamics of forest communities are intricately shaped by the ecological attributes of their surroundings, encompassing factors such as the features of the environment, the variety and abundance of species, the overall diversity, the spatial distribution of organisms, and regeneration status of species.13,14 Community structure is directly influenced by population structure, fundamental component of community. Consequently, development trend of community is clearly evident.
Regeneration is an essential component of forest management, as it is predictable by the structure of the population and ensures the desired species composition and replenishment. Successful regeneration of forest species is indicated by the population structure, which is defined by presence of adequate populations of saplings, seedlings, and young trees.15 The regeneration profile of a plant species' seedlings, saplings, and trees might be utilised to ascertain its possibilities for regeneration via the patterns of population dynamics.16
Himalayan forests are experiencing a variety of changes in their structure, density, composition, regeneration as a result of progressive biotic pressures that have been induced by uncontrolled lopping and harvesting of trees for fodder, fuel, and grazing.17,18
Dynamics of forest communities are significantly influenced by these biotic pressures, which also regulate19 regenerative capacity of a species. Vigour and rate of seed production of trees are reduced as a result of biotic interference, particularly the lopping of trees, which ultimately affects their regeneration status. A comprehensive understanding of forest dynamics is essential for conserving plant diversity.20 Earlier studies have indicated that Quercus lanata is found in scattered patches, often forming populations with individuals of varying sizes and ages, reflecting an uneven-aged structure with some large-sized trees. However, there is a need for updated and detailed information on its diversity status and population structure. Understanding the natural variation within populations is essential, as it supports species' ability to adapt, survive, and evolve over time. This becomes especially important for a species with significant ecological and economic value. If the current rate of decline continues, the species may face serious threats in the future. Key environmental factors encompassing soil characteristics, altitude, and seasonal rainfall—particularly during warmest, driest, and coldest quarters—as well as slope, have a strong influence on distribution of Q. lanata in Kumaun region. Present research focuses on evaluating forest composition, regeneration patterns, and population structure of Q. lanata. As Q. lanata is restricted to small patches, the study is significant to assess the future species composition in these forests.
Materials and Methods
Study area
The Indian Himalayan Region encompasses two mega-floristic zones: Garhwal and Kumaon. Uttarakhand Himalaya is one of them. The study area is situated in Nainital and Pithoragarh district of Kumaun Himalaya Uttarakhand. The study area is located between 29° 25’30” N latitude and 79°25’60” E longitude between 2050 –2200m elevations in Nainital district and 29° 30’54” N latitude and 80°14’31” E longitude between 1950 –2200m elevations in Pithoragarh district of Kumaun Himalaya. This region is characterized by temperate forests dominated by tree species such as oaks (Quercus spp.), R. arboreum, and Lyonia ovalifolia. Winter (November to February), summer (April to mid-June), and rainy (mid-June to September) are three primary seasons of temperate monsoon climate found at research locations. In addition to typical annual rainfall of 130.92mm and mean monthly rainfall of 3.7 to 451.38mm, mean lowest temperature ranged 5 - 21°C, and mean maximum temperature ranged from 17 - 28°C.21
Table 1: Description of study sites
Sites | Elevation (m) | Latitude | Longitude | Major species |
Nainital (Kilbury) | 2050-2200 | 29° 25'30'' | 79°25'60'' | Quercus lanata,Rhododendron arboreum, Quercus leucotrichophora |
Pithoragarh (Thalkedar) | 1950-2200 | 29° 30'54'' | 80°14'31'' | Quercus lanata, Lyonia ovalifolia, Rhododendron arboreum |
Sampling procedure
At each forest site, vegetation was analysed using the quadrat technique. 20 quadrats of 10 ×10m were laid randomly in each forest site for trees, 5 ×5m for saplings and 1 ×1m for seedlings. Sampling size and the number of quadrats were determined, and parameters such as density, total basal area (TBA), and Importance Value Index (IVI) were estimated following the methods described by Misra and Curtis and McIntosh.24,25
For developing population structure circumference at breast height (cbh), 1.37 m from the base of the tree with a meter tape was used. The data were categorized into eight circumference classes and population structure developed following i.e., seedlings(<10cm), saplings (10-30), 31–60 cm, 61–90 cm, 91–120 cm, 121–150 cm, and >150 cm, following the method of Good and Good.26 Shannon-Wiener information index was employed to determine species diversity (H') of each vegetation layer.27
![]()
Here, Ni implies the number of individuals of a species, and N indicates total number of individuals of all species in that stand.
Concentration of dominance (Cd) was computed by Simpson’s Index28
![]()
Here, Ni implies the number of individuals of a species, and N represents total number of individuals of all species.
Results
A total of 7 trees species were recorded across both sites. At site -1 the total tree density was 665 ind ha-1. With Q. lanatahaving the maximum density of 525 ind ha-1. Total basal area at this site was 36.96m² ha-1Q. lanata was dominant species followed by R. arboretum C. torulosa, Q. floribunda and L. ovalifolia at site -1. At site -2 total density was 1115ind ha-1. with Q. lanatahaving a density of 560 ind ha-1 Total basal area at this site was 21.28m2ha-1.
The density of Q. lanata was more or less the same across both sites. The Importance Value Index (IVI) of Q. lanata was 139.05 at site - 1 and 222.40 at site -2 (Table.2).
Table 2: Tree vegetation analysis of Quercus lanata dominated forest in Kumaun Region
Site-1 | Site-2 | |||||
Species | D (indha-1) | TBA (m2ha-1) | IVI | D (ind ha-1) | TBA (m2ha-1) | IVI |
Quercus lanata | 525 | 36.23 | 222.40 | 560 | 21.28 | 139.05 |
Q. floribunda | 20 | 1.64 | 17.87 | 10 | 0.26 | 3.16 |
M.esculenta | - | - | - | 25 | 0.7 | 7.26 |
Cupressus torulosa | 50 | 1.24 | 29.50 | - | - | - |
Lyonia ovalifolia | 15 | 0.32 | 5.74 | 100 | 2.3 | 33.94 |
Rhododendron arboreum | 55 | 1.10 | 24.50 | 180 | 5.22 | 49.02 |
Q. leucotrichophora | - | - | - | 240 | 7.2 | 67.57 |
Total | 665 | 40.52 | 1115 | 36.96 | ||
D- Density, TBA-Total basal area, IVI- Importance value index
Table 3: Seedling, Sapling Density and Total Basal Area of Quercus lanata dominated forests in the Kumaun Region
Species | Site-1 | Site-2 | ||||
Seedlings | Saplings | Seedlings | Saplings | |||
D (ind ha-1) | D (ind ha-1) | TBA (m2ha-1) | D (ind ha-1) | D (ind ha-1) | TBA (m2ha-1) | |
Q. lanata | 80 | 20 | 0.080 | 455 | 10 | 0.058 |
Q. floribunda | 10 | 10 | 0.028 | 10 | - | - |
C. torulosa | 10 | 5 | 0.033 | - | - | - |
R.arboreum | 15 | 65 | 0.023 | 55 | 5 | 0.025 |
Q.leucotrichophora | 15 | 5 | 0.260 | 130 | 50 | 0.224 |
M.esculanta | - | 5 | 0.023 | 190 | 20 | 0.098 |
L.ambrosa | - | 5 | 0.014 | - | - | - |
L.ovalifolia | - | 5 | 0.010 | 325 | 10 | 0.04 |
Total | 130 | 120 | 0.470 | 1165 | 95 | 0.444 |
D- Density, TBA-Total basal area
The total seedling density was 130 ind ha-1. at site-1 whereas site-2 it was 1165 ind ha-1 and the total sapling density was 120 ind ha-1 at site-1 and 95 ind ha-1 for site-2. Seedling density of Q. lanata ranged from 80 - 455ind ha-1, while sapling density ranged 10 - 20 ind ha-1 across both sites. Total basal area of Q. lanata saplings varied between 0.058 - 0.080 m² ha-1 (Table.3) across the sites. At site-1 the maximum sapling density was of R. arboreum and in site-2 of Q. leucotrichophora. Q. lanata, seedling density was higher at site-2, but sapling density was greater at site-1.
Tree Diversity index of study sites
The species diversity (H) for the tree layer across both sites ranged from 0.861 to 1.897. The diversity for saplings and seedlings varied between 1.868 and 2.164, and 1.719 and 2.09, respectively. Cd for tree layer ranged from 0.333 to 0.637, while for saplings it varied between 0.337 and 0.346, and for seedlings, it ranged from 0.272 to 0.417 (Table.4).
Table 4: Tree Diversity index of study sites
Site-1 | Tree | Sapling | Seedling |
H | 0.861 | 2.164 | 1.719 |
Cd | 0.637 | 0.337 | 0.417 |
Site-2 | Tree | Sapling | Seedling |
H | 1.897 | 1.868 | 2.09 |
Cd | 0.333 | 0.346 | 0.272 |
H- Diversity, Cd- Concentration of dominance
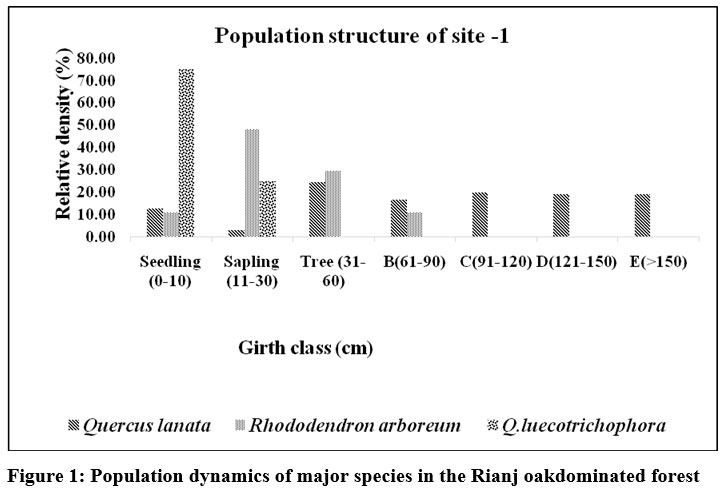 | Figure 1: Population dynamics of major species in the Rianj oakdominated forest
|
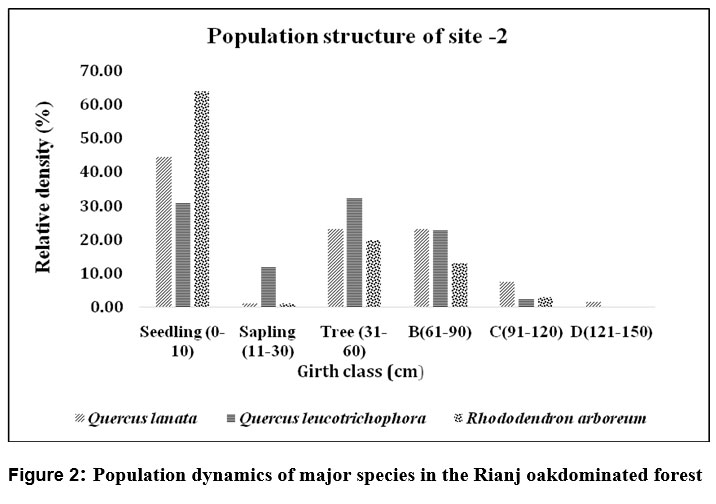 | Figure 2: Population dynamics of major species in the Rianj oakdominated forest
|
Population Structure
At site -1 Q. lanatashowes all girth classes from seedling (0–10 cm) to the highest girth class (>150 cm).Q. leucotrichophora is only represented in the seedling (0–10 cm) and sapling (11–30 cm) classes,with no individuals observed in higher girth classes, indicating poor recruitment into mature stages.R. arboreumis present in various girth classes, mainly in the sapling (11–30 cm) and small tree (31–60 cm), but is absent in the larger girth classes (>90 cm), indicating limited transition to mature stages fig1.Several ecological and anthropogenic disturbances such as logging, fuelwood collection that selectively removed mature trees, leading to a gap in older age classes. At site-1 tree density pattern generally decreases with increasing girth, which highlights the absence of older and larger trees.
At site-2 as the girth class increases (B 61 - 90cm, C 91-120cm, D121-150cm), the relative density decreases, witha limited number of species represented in the larger girth classes. Q. lanata shows fair regeneration status, as indicated by presence of considerable proportion of seedlings, mature trees. However, comparatively low density of saplings indicates there generation is limited during the sapling stage conversion rate from seedling to sapling was poor. The absence of trees in the highest girth class (D) indicates a lack of very old or mature trees in the site fig2. The absence of higher girth class trees may be due to low recruitment rates, high mortality, other disturbances such as logging or grazing, or environmental stress. The restricted conversion of seedlings of Q. lanata to sapling stage shows that the species is facing regeneration problem.
Discussion
Density of saplings and seedlings is criterion implemented for assessing regeneration status of tree species in any forest. A population's reproductive status and future outcome are determined by the ratio of various age groups.29 Regeneration maintains forest vitality by ensuring a continuous replacement of aging trees with saplings and seedlings.30 In space and time, forest's wealth is contingent upon potential regenerative status of species that comprise the forest stand.31
In the present study across both the sites the tree density of Q. lanata ranged between 525 and 560 ind ha-1. which was lower than the density reported by Singh et al.,32 for the species Dhar et,al,.33 The structural diversity and representativeness of forest vegetation in the Kumaun Himalaya were studied, and the density of Q. lanata was reported at 832 ind ha-1.
Sapling density of Q. lanata across both the sites was 10 and 20ind ha-1. In oak-dominated forests of Nainital, sapling density ranges from 500 to 950 ind ha-1 as reported by Singh et.,al.34, while in the Binsar Wildlife Sanctuary, Almora, total sapling density ranged from 20 to 380 ind ha-1 as reported by Khan and Arya.35 These values indicate that the sapling density observed in this study is lower than that reported previously (Table 3).
Seedling density of Q. lanata at site 1 has been 80 ind/ha and at site 2 density was 455 ind ha-1. Seedling density at site1 has been lower than that at site 2. The low seedling density at site 1 could be due to low seed production, failure of germination, dense canopy cover, herbivory, and anthropogenic disturbances. Drywood collection is common in the area in which the study is located.
In the present study, total seedling density varied between 130 and 1165 ind ha-1 across both sites of Kumaun Himalaya. Similarly, a previous study reported by Khan and Arya.35 the seedling density ranges from 640 to 1880 ind ha-1 in the central Himalaya, which was higher than the value of present study in oak-dominated forests.
Total basal region of Q. lanata in the current study ranges between 21.28 and 36.23 m2ha-1across both the sites. Which was lower than the total basal area reported by Singh SP, Singh JS.9 Total basal area of Q. lanata in Kumaun Himalaya 67.7 m2ha-1 33 for the species. In current investigation, total basal area of Q. lanata dominated forest ranged from 36.96 to 40.52 m²/ha in Kumaun Himalaya.
Q. lanata, ecologically and economically essential oak species in Kumaun Himalaya, exhibits a highly restricted distribution, typically forming small, fragmented forests. Q. lanata distribution in Kumaun region is strongly influenced by several environmental factors, including altitude, soil characteristics, slope, and seasonal precipitation. While altitude plays a key role, rainfall emerges as a critical factor for the species regeneration, as acorn germination typically occurs under moist conditions during and shortly after the monsoon season. However, once the monsoon ends, field conditions often become dry, which can threaten the survival of the germinated seedlings. Therefore, an extended dry period following the pre monsoon drought season may significantly reduce the survival rate of Q. lanata seedlings. Uncontrolled grazing by domestic livestock has a detrimental impact on regeneration of forests, as it removes juvenile seedlings and saplings, leading to soil loss because of trampling.36 The seedling regeneration status of studied species was low. The seedling density of all the other associated species was high but conversion of seedling into sapling was relatively poor. Poor regeneration of species in Himalayan mountain forests has been reported by other workers from time to time.37 Earlierstudies by Bisht S, et. al.38 have show fair regeneration of Q.lanata in protected Sanctuary. The seedling density in the study was 245 ind ha-1. It is evident that the species is regenerating in protected area where anthropogenic pressure is low.
The diversity of the Q. lanata-dominated forest at Site 1 was recorded as 0.861, which is lower than the value of 1.88 reported by Dhar et.al. 33.In current research, the diversity (H) for tree species was 0.861 - 1.89, while for saplings and seedlings, the values ranged from 0.337 - 2.164 and 0.272 - 2.09 (Table.4) respectively, across both the site34 reported diversity values ranging from 1.31 - 2.69 for trees, 1.78 - 1.84 for saplings, and 0.87 - 1.50 for seedlings in an oak-dominated forest in Kumaun Himalaya. Diversity indices for saplings and seedlings were comparatively higher than those presented in the earlier study.
Q. lanata is capable of regenerating even under closed canopy conditions, and it occasionally coexists with other species, sharing the forest canopy. Since it is widely used for fuelwood, fodder, and leaf litter, local communities often engage in selective extraction or lopping of these trees. This practice leads to canopy openings, which opens up the canopy and facilitates regeneration.39
Tree size class distribution is the primary indicator of forest population structure.23,15 The population structure of site 1 and site 2 shows notable contrasts in girth class distribution among various species. In site-1, the relative density of Q. lanata is higher in the seedling (0-10 cm) and sapling (11-30cm) classes. Therefore, density decreases in larger girth classes, suggesting fewer mature individuals. Site 2 show highest relative density is observed in the seedling class (0–10 cm), indicating active recruitment but the conversion of seedling into sapling was poor. The limited representation of higher girth class trees at both sites may be due to factors such as low recruitment rates, high mortality, or disturbances like grazing, logging, or environmental stress. The absence of older trees also indicates that the forests might be in an earlier successional stage, where mature individuals are not yet established. Site 2 shows a more sustainable population structure with consistent presence across various girth classes, whereas site1 appears to depend largely on younger individuals, which might result in challenges in maintaining a stable number of mature individuals. The lack of mature trees in the larger girth classes observed in this study likely reflects the effects of past selective logging or elevated mortality rates caused by anthropogenic disturbances. In addition, climate variability such as irregular rainfall and rising temperature these problems by disrupting the processes of seed development and germination.40
Table 5: Comparison of tree, sapling, seedling density and regeneration status of Q. lanata with earlier studies in oaks of central Himalayas
Sl. no | Species | Tree density (ind/ha) | Sapling density (ind/ha) | Seedling density (ind/ha) | Regeneration status | Sources |
1 | Q. lanata | 672 | Absent | Absent | Poor | Singh and Singh, 19869 |
2 | Q. lanata | 993 | Absent | Absent | Poor | Singh et al., 199432 |
3 | Q. floribunda | 760 | Absent | Absent | Poor | Singh et al., 201434 |
4 | Q. lanata | 570 | 900 | 933 | Fair | Saxena and Singh 198423 |
5 | Q. leucotrichophora | 210 | 60 | 40 | Poor | Khan and Arya 201735 |
6 | Q. floribunda | 583 | 50 | 189 | Poor | Giri et al., 2008 |
7 | Q. semecarpifolia | 142.25 | 232.7 | 52.65 | Poor | Singh and Rawat 20127 |
8 | Q. floribunda | 740 | 260 | 70 | Fair | Lodhiyal et al., 2015 |
9 | Q. lanata | 1000 | 480 | 1840 | Fair | Bisht et al.,202138 |
10 | Q. lanata | 525 | 20 | 80 | Poor | Present study site1 |
11 | Q. lanata | 560 | 10 | 455 | Poor | Present study site2 |
Conclusion
The current study strongly indicate that some degree of protection is required for proper regeneration of Q lanata in the Himalayan region where it is exists in very small pockets.More studies in the Himalayan region are required for a clear assessment of its regeneration potential and survival. In the Himalayan region poor to fair regeneration with excessively lower conversion of seedling into sapling is a matter of huge concern regarding survival of the species in the coming future. More conservation efforts are required.
Acknowledgement
The author would like to thank Uttarakhand Forest Department for granting permission to work in the forest area. Author are Thankful to Head, Department of Forestry, Kumaun University, Nainital for providing necessary resources and support for this study.
Funding Sources
The author(s) received no financial support for the research, authorship, and/or publication of this article.
Conflict of Interest
The authors do not have any conflict of interest.
Data Availability Statement
The data support the findings of study is available from the corresponding author upon reasonable request
Ethics Statement
This research did not involve human participants, animal subjects, or any material that requires ethical approval.
Informed Consent Statement
This study did not involve human participants, and therefore, informed consent was not required.
Permission to Reproduce Material from other sources
Not applicable
Author Contributions
Geeta Sharma: Conceptualization, Methodology, Field Investigation, Data Collection, Data Analysis, Writing original Draft.
Ashish Tewari: Data Analysis, Writing – Review & Editing.
Shruti Shah: Writing – Review & Editing, Proofreading, Quality Check.
References
- Joshi AK, Juyal D. Traditional and ethnobotanical uses of Quercus leucotrichophora A. Camus (Quercus oblongata D. Don) in Kumaun and Garhwal regions of Uttarakhand, India: a review. Int J Herb Med. 2017;5(5):6–8.
- Sati VP. Himalaya on the Threshold of Change. Springer; 2020.
CrossRef - Singh B, Bhatt BP, Prasad P. Effect of seed source and temperature on seed germination of Celtis australis L.: a promising agroforestry tree-crop of Central Himalaya. For Trees Livelihoods.2004;14:53–60.
CrossRef - Bhatt J, Tewari A, Mittal A. Regeneration problem in Quercus leucotrichophora A. Camus in Nainital Forest Division of Kumaun Himalaya. Int J Pure Appl Biosci. 2015;3(3):284–290.
- Kala CP. Studies on the indigenous knowledge, practices and traditional uses of forest products by human societies in Uttaranchal State of India. Almora: GB Pant Institute of Himalayan Environment and Development; 2004:82.
- Valdiya KS. Dynamic Himalaya. Hyderabad: University Press Ltd; 1998:178.
- Troup RS. Silviculture of Indian Trees. Vol I–III. Oxford: Clarendon Press; 1921.
- Singh G, Rawat GS. Depletion of oak (Quercus spp.) forests in the Western Himalaya: grazing, fuelwood and fodder collection. In: Okia CA, ed. Global Perspectives on Sustainable Forest Management. InTech; 2012:29–42.
CrossRef - Singh SP, Singh JS. Structure and function of the Central Himalayan oak forests. Proc Plant Sci.1986;96:159–189.
CrossRef - Khan M, Khan SM, Ilyas M, Alqarawi AA, Ahmad Z, Abd_Allah EF. Plant species and community assessment in interaction with edaphic and topographic factors: an ecological study of Mount Eelum, District Swat, Pakistan. Saudi J Biol Sci. 2017;24(4):778–786.
CrossRef - Tarin MW, Nizami SM, Jundong R, et. al. Range vegetation analysis of Kherimurat scrub forest, Pakistan. Int J Dev Sustain. 2017;6(10):1319-1333.
- Barnes BV, Zak DR, Denton SR, Spuit SH. Forest Ecology. New York, NY: John Wiley & Sons; 1998.
- Pandita S, Kumar V, Dutt HC, et. al. Environmental variables visàvis distribution of herbaceous tracheophytes on northern subslopes in Western Himalayan ecotone. Ecol Process. 2019;8(1):1-9. doi:10.1186/s137170190200x.
CrossRef - Joshi VC, Bisht D, Sundriyal RC, Pant H. Species richness, diversity, structure, and distribution patterns across dominant forest communities of low and mid hills in the Central Himalaya. Geol Ecol Landscapes. 2022;():1-11.
CrossRef - Khan ML, Rai JPN, Tripathi RS. Population structure of some tree species in disturbed and protected subtropical forests of northeast India. Acta Ecol Oecol Appl. 1987;():247-255.
- Teketay D. Seedling populations and regeneration of woody species in dry Afromontane forests of Ethiopia. For Ecol Manage. 1997;98(2):149-165.
CrossRef - Bargali SS, Rana BS, Rikhari HC, Singh RP. Population structure of Central Himalayan blue pine (Pinus wallichiana) forest. Environ Ecol.1989;7:431-436.
- Kumar M, Sharma CM, Rajwar GS. Community structure and diversity of a subtropical forest of Garhwal Himalaya. Indian For. 2004;130(2):207-214.
- Pickett STA, White PS, eds. The Ecology of Natural Disturbance and Patch Dynamics. New York, NY: Academic Press; 1985.
- Murali KS, Shankar U, Shankar RU, Ganeshiah KN, Bawa KS. Extraction of nontimber forest products in the forests of Biligiri Rangan Hills, India: impact on regeneration, population structure and species composition. Econ Bot.1996;50:252-269.
CrossRef - Fartyal A, Bhatt AB, Lodhiyal LS, et. al. Altitudinal variation in plant community, population structure, and carbon stock of Quercus semecarpifolia (Sm.) forests in Kumaun Himalaya. J Environ Biol. 2022;43(1):133-146.
CrossRef - Saxena AK, Singh JS. A phytosociological analysis of woody species in forest communities of a part of Kumaun Himalaya. Vegetatio.1982;50:3-22.
CrossRef - Saxena AK, Singh JS. Tree population structure of certain Himalayan forest associations and implications concerning their future composition. Vegetatio. 1984;58(2):61-69.
CrossRef - Misra R. Ecology Workbook. Calcutta, India: Oxford & IBH Publishing; 1968.
- Curtis JT, McIntosh RP. The interrelationship of certain analytic and synthetic phytosociological characters. Ecology.1950;31:438-455.
CrossRef - Good NF, Good RE. Population dynamics of tree seedlings and saplings in mature eastern hardwood forest. Bull Torrey Bot Club.1972;99:172-178. doi:10.2307/2484571.
CrossRef - Shannon CE, Weaver W. The Mathematical Theory of Communication. Urbana, IL: University of Illinois Press; 1963.
- Simpson EH. Measurement of diversity. Nature.1949;163:688.
CrossRef - Odum EP. Fundamentals of Ecology. Philadelphia, PA: W.B. Saunders; 1971.
- Malik ZA, Bhatt AB. Regeneration status of tree species and survival of their seedlings in Kedarnath Wildlife Sanctuary and its adjoining areas in Western Himalaya, India. Trop Ecol. 2016;57(4):677-690.
- Jones RH, Sharitz RR, Dixon PM, et. al. Woody plant regeneration in four floodplain forests. Ecol Monogr. 1994;64(3):345-367.
CrossRef - Singh SP, Adhikari BS, Zobel DB. Biomass, productivity, leaf longevity, and forest structure in the Central Himalaya. Ecol Monogr. 1994;64(4):401-421.
CrossRef - Dhar U, Rawal RS, Samant SS. Structural diversity and representativeness of forest vegetation in a protected area of Kumaun Himalaya, India: implications for conservation. Biodivers Conserv. 1997;6:1045-1062.
CrossRef - Singh N, Tamta K, Tewari A, Ram J. Vegetational analysis and regeneration status of Pinus roxburghii Roxb. and Quercus leucotrichophora forests of Nainital Forest Division. Glob J Sci Front Res. 2014;14(3):41-47.
- Khan AH, Arya D. Analysis of forest vegetation in Binsar Wildlife Sanctuary, Kumaun Himalaya, Uttarakhand, India. Am Eurasian J Agric Environ Sci. 2017;17(4):336-342.
- Saberwal VK. Pastoral politics: Gaddi grazing, degradation, and biodiversity conservation in Himachal Pradesh, India. Conserv Biol. 1996;10(3):741-749.
CrossRef - Singh N, et.al. Tree regeneration pattern and size class distribution in anthropogenically disturbed sub alpine treeline areas of Indian Western Himalaya. Int J Sci Technol Res. 2019;8(8):537-546.
- Bisht S, et.al. Carbon pool in Quercus lanuginosa (D.Don) forest of Askot Wildlife Sanctuary, Western Himalaya in relation to stand attributes. Indian For. 2022;148(7):709-717.43
CrossRef - Wangda P, Ohsawa M. Structure and regeneration dynamics of dominant tree species along an altitudinal gradient on dry valley slopes of the Bhutan Himalaya. For Ecol Manage. 2006;230(1.3):136-150.
CrossRef Bhandari MS, Meena RK, Shankhwar R, et. al. Global warming scenario depicts enhanced spatial distribution of Quercus lanata in the western Himalayas. Int J Global Warming. 2020;22(3):255-271.
CrossRef







