Optimization of Cellulases under Solid State Fermentation by Newly Isolated Fungus: An Environmentally Sustainable Approach
Anil Kumar1
 , Meenakshi Suhag2
*
, Naveeta Dhaka1
, Joginder Singh3
, Ritu Nandal1
and Rajesh Dhankhar4
, Meenakshi Suhag2
*
, Naveeta Dhaka1
, Joginder Singh3
, Ritu Nandal1
and Rajesh Dhankhar4
1
Department of Botany,
Pt. NRS Govt. College,
Rohtak,
Haryana
India
2
Institute of Environmental Studies,
Kurukshetra University,
Kurukshetra,
Haryana
India
3
laboratory of Environment Biotech,
Department of Botany,
AIJHM College,
Rohtak,
Haryana
India
4
Department of Environmental Science,
MD University,
Rohtak,
Haryana
India
http://dx.doi.org/10.12944/CWE.19.3.16
Copy the following to cite this article:
Kumar A, Suhag M, Dhaka N, Singh J, Nandal R, Dhankhar R. Optimization of Cellulases under Solid State Fermentation by Newly Isolated Fungus: An Environmentally Sustainable Approach. Curr World Environ 2024;19(3). DOI:http://dx.doi.org/10.12944/CWE.19.3.16
Copy the following to cite this URL:
Kumar A, Suhag M, Dhaka N, Singh J, Nandal R, Dhankhar R. Optimization of Cellulases under Solid State Fermentation by Newly Isolated Fungus: An Environmentally Sustainable Approach. Curr World Environ 2024;19(3).
Download article (pdf)
Citation Manager
Publish History

Introduction
Bioethanol, derived from lignocellulosic biomass (LB), possesses the potential to substitute our reliance on conventional petroleum fuel. LB, being a renewable and carbon-neutral resource, serves as an excellent raw material with vast potential for sustainable bioethanol production. This, in turn, can contribute to mitigating climate change and ensuring long-term energy security. The bioconversion process of lignocellulosic biomass (LB) into ethanol involves three main steps: pretreatment, saccharification/hydrolysis, and fermentation. Cellulases play a crucial role in generating fermentable sugars for ethanol production. Consequently, a significant quantity of cellulase enzymes is required to scale up the conversion process to an industrial level. However, the high production cost of commercially available cellulases makes the entire process expensive. In recent years, numerous studies have focused on exploring new approaches to develop a cost-effective enzyme production process 1. In this particular context, extensive research has been executed on various microorganisms, particularly filamentous fungi such as Aspergillus niger 2,3, Trichoderma sp.4,5, and Penicillium sp.6, to explore their capacity for producing cellulases in high quantities. Additionally, strain improvement using genetic engineering and recombinant DNA technologies has also been investigated to enhance cellulases production. These efforts hold significant potential for creating cost-effective and efficient enzyme production processes.
In recent years, remarkable inclination towards adopting solid-state fermentation instead of submerged fermentation for producing microbial enzymes, mainly because of its superior energy efficiency have witnessed 7. Several studies have compared the capabilities of SSF and SmF, consistently demonstrating that SSF yields better hydrolytic performance and higher initial reaction rates, resulting in higher levels of extracellular enzymes8,9. In SSF, microorganisms grow on the surface of solid substrates without access to free water. This enables faster and more cost-effective downstream processing with reduced purification and extraction requirements10 Additionally, SSF allows the utilization of low-cost agricultural waste or residues as substrates for enzyme production, thereby reducing production costs 11. Numerous reports have demonstrated successful cellulase production through SSF using a variety of substrates, including wheat bran 12, oil palm waste 13, agro-waste 14, waste paper 15, agricultural kitchen waste residue 16, Jatropha curcas seed cake 17, sweet sorghum stalk 18, sugarcane residue19, rice straw 20, apple pomace 21, corn straw22, and fruit wastes23, among others.
Fungi exhibit greater involvement in solid state fermentation (SSF) compared to bacteria due to the low water activity present in solid substrates24. In this study, various fungal isolates obtained from soil samples were investigated, and the fungal isolate displaying the highest hydrolytic potential was selected for SSF using two cost-effective substrates: municipal solid waste (MSW) and water hyacinth (Eichhornia crassipes) to study cellulases production. Municipal solid waste, which contains a high content of lignocellulosic material, was chosen as a promising substrate that has received limited attention in cellulase production studies. On the other hand, water hyacinth was selected due to its rapid and problematic growth in water-bodies, causing environmental issues25, but containing high cellulose and low lignin content, with approximately 60% cellulose, 8% hemicellulose, and 17% lignin which makes it a suitable and cost-effective substrate for bioethanol production26.
Materials and Methods
Isolation and screening of microorganisms
Fungi capable of producing cellulase were isolated from soil samples collected in the districts of Kurukshetra, Jind, Kaithal, Ambala, Rohtak, and Jhajjar in Haryana, India (27°37 to 30°35N and 74°28 to 77°36E). The isolation process involved serial dilution of the soil samples followed by culturing on Potato Dextrose Agar (PDA) media27. Each fungal isolate was spot inoculated on modified Mandel's and Sternburg (MS) basal agar medium28 (composition: Proteose Peptone - 1.4g/l, KH2PO4 - 1gm/l, (NH4)2SO4 - 2.0gm/l, MgSO4.7H2O - 0.3gm/l, Urea - 0.3gm/l, , FeSO4.7H2O - 0.005gm/l, CaCl2 - 0.3gm/l, MnSO4.H2O - 0.0016gm/l, ZnSO4.7H2O - 0.0014gm/l, CoCl2 - 0.002gm/l) at pH 5.0, supplemented with 1% (w/v) CMC (Carboxymethylcellulose) in petriplates. The fungal cultures were then incubated at 30ºC for three days. Following incubation, every culture plate was exposed to a Congo red solution (0.1%) for a duration of 15 min., proceeded by rinsing with 1M sodium chloride to eliminate any surplus dye. The measurement of the diameter of the hydrolysis zone surrounding the fungal colonies served as an indicator of cellulase activity.
Fungal isolates displaying larger hydrolysis zones were chosen for cellulase enzyme production. The screening of cellulase production through SSF was conducted using the untreated and dried lignocellulosic biomasses of E. crassipes as a carbon source. SSF was conducted in 500 mL Erlenmeyer flasks containing 5g of dried Eichhornia crassipes biomass with particle sizes ranging from 0.2 to 1.0 mm. To initiate SSF, the biomass was moistened first with 30 mL of MS basal medium and then autoclaved to ensure sterility for 20 minutes at 121°C. A 1.0 mL suspension containing 5x107 spores was used to inoculate the sterilized flasks, which were incubated at 30°C with intermittent shaking. On the 4th day, the culture flasks were removed, and the enzymes were extracted by adding 10 mL of Citrate Buffer (0.05 M, with a pH of 4.8) per gram of substrate. The mixture was shaken on an incubator shaker with a rotation of 200 per minute (rpm), kept at a constant temperature (30°C) for an interval of one hour, and then filtered. The filtrate was then carefully subjected to centrifugation at a speed of 10,000 revolutions per minute for a duration of 10 minutes with a temperature of 4°C, which successfully yielded a vibrant supernatant. Subsequently, the cellulase activity of this supernatant solution was assayed and compared to that of Trichoderma reesei NCIM 992, a commercially available fungus strain. The reference strain was sourced from the “National Collection Centre of Industrial Microorganisms” located in Pune, India.
Identification of potent cellulolytic fungal isolates
The fungal isolate-130, analyzed for its strong cellulase production, underwent primary identification at the Institute of Microbial Technology (IMTECH) in Chandigarh. This identification was achieved through the analysis of the Translation Elongation Factor gene sequence. The obtained sequence was compared with the existing database sequences using a “Basic Local Alignment Search Tool (BLAST)” to examine its resemblance with the known one. For a more in-depth examination of its evolutionary relationships, a comprehensive phylogenetic tree was created using the “Mega 5 software”.
Lignocellulosic substrate
The dried and untreated lignocellulosic biomasses (LBs) of E. crassipes and Municipal Solid Waste (MSW) of particle sizes 0.2 to 1.0 mm were used as substrates for solid-state fermentation.
Preparation of inoculum
The fungal slants were prepared by dispensing a sterile solution of normal saline mixed with 0.1% (v/v) Tween-80 into each tube, with volumes ranging from 20 to 40 mL. Using a sterile loop, the spores were gently dislodged from the fungal culture and introduced into the saline solution. To ensure an even distribution of spores, the liquid was vigorously shaken. For a precise determination of spore concentration, a hemocytometer was employed, and 1 mL aliquot of the suspension estimated to contain approximately 1x107 spores was utilized for spore count procedure.
Solid state fermentation
In the experiment, Erlenmeyer flasks with volumes of 250mL and 500mL were used. The flask of 250mL contained 5g of municipal solid waste (MSW), while the 500mL flask contained 5g of Eichhornia crassipes biomass. These materials were combined with a specific quantity of Mandels and Sternburg (MS) basal medium29, which consisted of the following components per liter: protease peptone-1g, (NH4)2SO4-1.4gm, KH2PO4-2.0gm, Urea-0.3gm, MgSO4 .7H2O - 0.3gm, CaCl2-0.3gm, FeSO4.7H2O - 0.005gm, , ZnSO4.7H2O-0.0014gm, CoCl2 -0.002gm, MnSO4.H2O-0.0016gm and Tween-80 (0.1% v/v). Additionally, glucose (0.1% w/v) was supplemented into the medium. The entire mixture was sterilized using an autoclave (at 121°C) for 20 minutes before inoculation with 1.0mL of a suspension containing 5x107 spores. To sustain the cultures, all flasks then were incubated with periodic shaking (at 30°C temperature). At predetermined intervals, the culture was removed for cellulase activity analysis, focusing on the measurement of three enzymes: FPase (filter paperase), CMCase (carboxymethylcellulase), and BGL (B-glucosidase).
Enzyme extraction
To extract the contents of the flasks, a substrate-to-citrate buffer ratio of 1:20 was employed, with the buffer consisting of a 0.05M solution at pH 4.8. The flasks were placed in an incubator and shaken for one hour at 200 rpm (30°C temperature). Afterward, the entire mixture was filtered to separate the solid and liquid components using a muslin cloth. To eliminate any remaining spores, the obtained filtrate underwent 10,000 rpm centrifugation at 4°C for 15 minutes. The subsequent clear supernatant obtained was then used for further experiments.
Enzyme assays
To calculate FPase activity, we used a conversion factor indicating 0.37 FPUs of enzyme yields 2.0mg of glucose when assessed during the specified assay conditions30. For estimating CMCase activity (U/mL), we utilized the fact that 0.185 units of enzymes release 0.5mg of glucose during the established conditions30. The assessment of BGL activity was based on the utilization of P-Nitrophenyl-D-Glucopyranoside (PNPG) as a substrate31. In the context of SSF, the quantity of B-glucosidase was quantified in units per gram of dry substrate (U/g). Here, 1 U represented the specific volume of enzyme needed to produce one mole of p-Nitrophenol in one minute, provided that the enzyme is operated under standardized assay conditions.
Optimization of fermentation conditions in solid state
The investigation into cellulase production involved assessing the impact of various factors using an OFAT (One-Factor-At-A-Time) design. The factors under scrutiny included moisture content, incubation time, pH, and nitrogen sources. To determine the impact of initial moisture level on cellulase synthesis, the substrate-to-media ratio (w/v) was modified from 1:1 to 1:3 for municipal solid waste (MSW) and from 1:4 to 1:7 for Eichhornia crassipes. Further analysis of cellulase production was conducted at the optimized substrate moisture ratios of 1:2 for MSW and 1:6 for Eichhornia crassipes. The fermentation process spanned a duration of 7 days, with cellulase production monitored at 24-hour intervals from the 3rd to the 7th day. To examine the influence of the initial pH of the culture on cellulase activity during the optimal incubation time, the pH of the MS medium was regulated by introducing 1N HCl or 1N NaOH, covering a range of 4.0 to 8.0. Additionally, the impact of diverse nitrogen sources on cellulase production was investigated using a complex MS medium. The medium incorporated varying amounts and combinations (I to VI) of nitrogen sources of both organic and inorganic origin, maintaining the optimal incubation period and starting pH of the moistening medium.
0.1% PP + 0.03% Urea + 0.14% AS
0.05% PP + 0.03% Urea+ 0.14% AS
0.1% PP+ 0.03% Urea + 0.28% AS
0.1% PP+ 0.03% Urea + 0.05% YE + 0.21% AS
0.05% PP + 0.03% Urea+ 0.1% YE + 0.21% AS
0.2% PP+ 0.03% Urea + 0.28% AS
AS = Ammonium Sulphate, PP = Protease Peptone, YE = Yeast Extract
Statistical tool
All experiments were conducted in triplicate to confirm the accuracy of the results. The experiments data were analyzed and presented as mean value accompanied by the standard error of the mean to give a clearer understanding of the variability. Calculation of the statistical analyses were performed using MS Excel.
Results
Identification of Fungal Strains
In this comprehensive study, a remarkable collection of 199 fungal isolates was meticulously procured from a diverse array of 50 soil samples spanning various regions of Haryana. To ascertain their cellulase production capabilities, a meticulous qualitative screening was carried out employing the plate assay method. Remarkably, out of the myriad fungal isolates, a select group of eight strains (isolates 2, 7, 44, 55, 81, 87, 130, and 148) exhibited significantly augmented zones of hydrolysis, thereby indicating a promising cellulase activity (Table 1). These exceptional fungal isolates, which displayed immense potential, were subsequently subjected to a rigorous secondary screening process aimed at assessing cellulase production under solid-state fermentation (SSF) conditions.
Table 1: Hydrolytic zones of different fungal isolates
Name/ No. of Fungal Isolates | Zone of hydrolysis (cm) | Name/ No. of Fungal Isolates | Zone of hydrolysis (cm) |
2 | 9.0±0.3 | 81 | 8.8±0.2 |
7 | 8.7±0.3 | 87 | 9.3±0.3 |
44 | 8.8±0.2 | 130 | 9.4±0.2 |
55 | 9.1±0.2 | 148 | 8.7±0.3 |
A comparative analysis of cellulase production under SSF was performed, pitting selected fungal isolates against the commercial strain Trichoderma reesei NCIM 992 (Figure 1). Remarkably, fungal isolates 55, 81, 87, 130, and 148 displayed significantly higher activity levels of FPase, CMCase, and BGL in comparison to T. reesei NCIM 992. Conversely, fungal isolates 2, 7, and 44 exhibited lower production of FPase and CMCase, although their BGL activities were mostly comparable, except for fungal isolate 2, which showcased higher B-glucosidase activity than T. reesei NCIM 992. Among all the fungal isolates, isolate 130 exhibited the maximum values of FPase, CMCase, and BGL, measuring at 38.57, 103.49, and 23.08 U/g, respectively, in contrast to 28.91, 82.51, and 6.11 U/g observed in Trichoderma reesei NCIM 992. Due to its exceptional cellulase activity, fungal isolate-130 was chosen as the prime candidate for subsequent trials among all the fungal isolates.
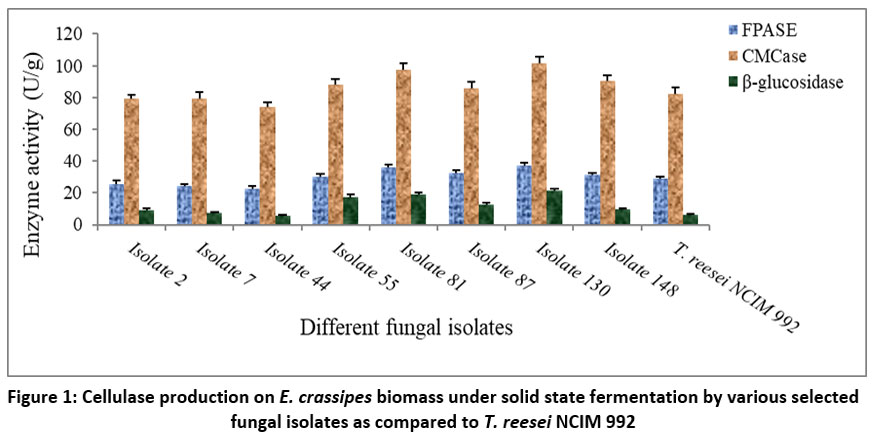 | Figure 1: Cellulase production on E. crassipes biomass under solid state fermentation by various selected fungal isolates as compared to T. reesei NCIM 992
|
Identification of Fungal Isolate
Based on an initial microscopic examination of morphological features, the fungus in question was identified as a member of the Trichoderma genus. To further confirm the molecular identity of fungal isolate-130, 19 nucleotide sequences for Translation Elongation Factors were utilized and subjected to rigorous analysis. To ensure robust and accurate identification, a phylogenetic tree was constructed using the Kimura-2-parameter (K2P) model, and the gene sequence database and MEGA 5 software were employed (Figure 2). Notably, the selected strain exhibited a close affiliation with Trichoderma atroviride (AF456891), a previously reported deuteromycetes fungus, as evident from their shared clade. Fusarium tricinctum (AB674263) served as the outgroup species. Consequently, the chosen strain was officially designated as T. atroviride AD-130, as per GenBank Accession KJ997913, and subsequently employed in solid state fermentation experiments, utilizing E. crassipes and municipal solid waste (MSW) as substrates, to explore its potential for producing desired enzymes.
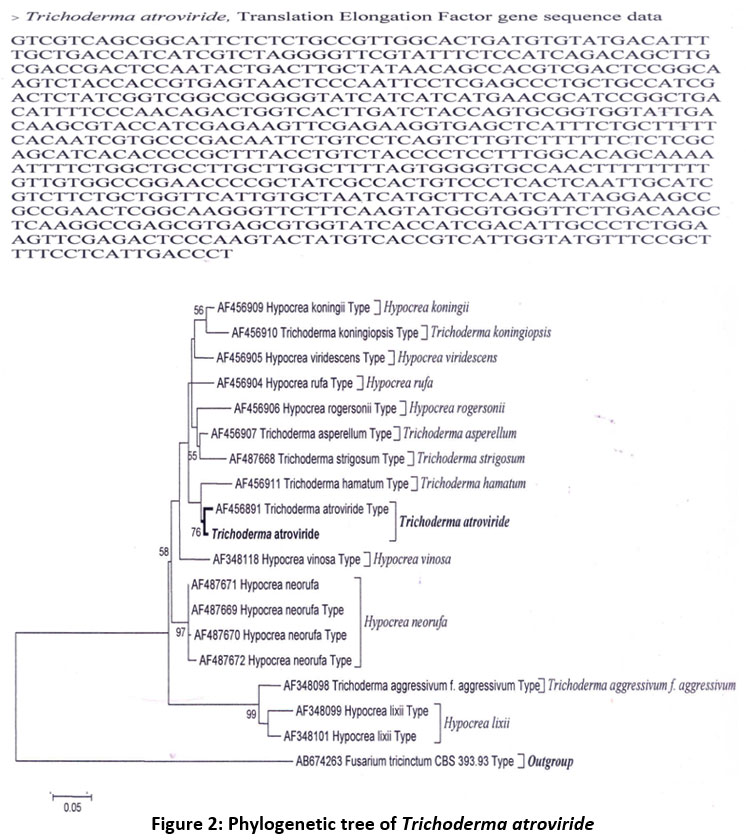 | Figure 2: Phylogenetic tree of Trichoderma atroviride
|
Impact of Moisture Level on Cellulase yield
In the present work, the effect of moisture level on enzyme activity was examined at the 5.0. pH of the fermentation medium while setting the temperature at 30°C. The enzyme activities were measured on the 5th day of fermentation. Figures 3 and 4 depict the impact of the primary moisture level on cellulase yields by T. atroviride AD-130 cultivated through SSF, using E. crassipes biomass and MSW residue as substrates, respectively. When E. crassipes was utilized as the substrate in SSF, the maximum activity of FPase was recorded at 41.17U/g, followed by CMCase at 141.83U/g, and BGL at 25.16U/g, all achieved at a substrate-moisture ratio of 1:6. On the other hand when MSW was employed as the substrate, the maximum production of all enzymes was found at a substrate to moisture ratio of 1:2, with FPase reaching 42.12U/g, CMCase at 79.54U/g, and B-glucosidase at 39.83U/g. However, it should be noted that a reduction in cellulase activity was observed in all other substrate-moisture ratios. Consequently, further investigations were carried out to assess the enzyme production capabilities of T. atroviride AD-130 through solid-state fermentation utilizing E. crassipes and MSW as carbon sources with the substrate-to-moisture ratios of 1:6 and 1:2, respectively.
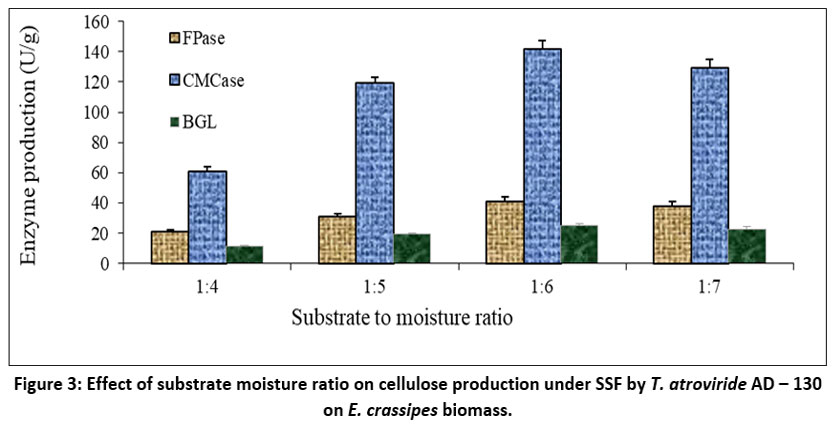 | Figure 3: Effect of substrate moisture ratio on cellulose production under SSF by T. atroviride AD – 130 on E. crassipes biomass.
|
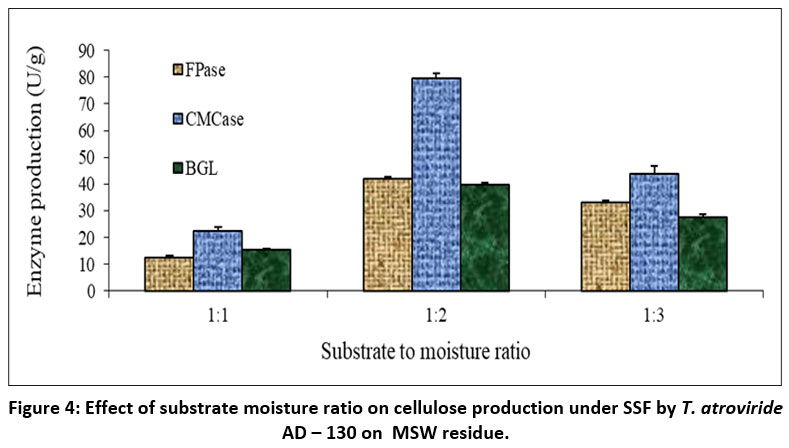 | Figure 4: Effect of substrate moisture ratio on cellulose production under SSF by T. atroviride AD – 130 on MSW residue.
|
Impact of Incubation Period on Cellulase Production
The impact of incubation time, from 3 days to 7 days, on fungal cellulases production was investigated using T. atroviride AD-130 under SSF with E. crassipes and MSW as carbon sources in substrate to moisture ratio of 1:6 and 1:2, respectively. The results, illustrated in Figure 5, exhibited that the highest enzyme production occurred on the 5th day. Maximum yields of FPase, CMCase, and BGL were achieved with MSW: 42.12U/g, 79.54U/g, and 39.83U/g, respectively. On the other hand, E. crassipes demonstrated maximum yields of FPase (41.17U/g), CMCase (141.83U/g), and BGL (25.16U/g). Notably, a comparison between the 3rd and 7th day revealed significant differences in enzyme production. On the 3rd day, the activities were 18.96U/g, 51.87U/g, and 12.96U/g for FPase, CMCase, and BGL on E. crassipes, respectively, and 19.32U/g, 26.87U/g, and 14.53U/g on MSW, respectively. However, on the 7th day, higher enzyme activities were observed: 34.11U/g, 121.71U/g, and 19.56U/g for FPase, CMCase, and BGL on E. crassipes, and 35.16U/g, 65.27U/g, and 31.14U/g on MSW, respectively.
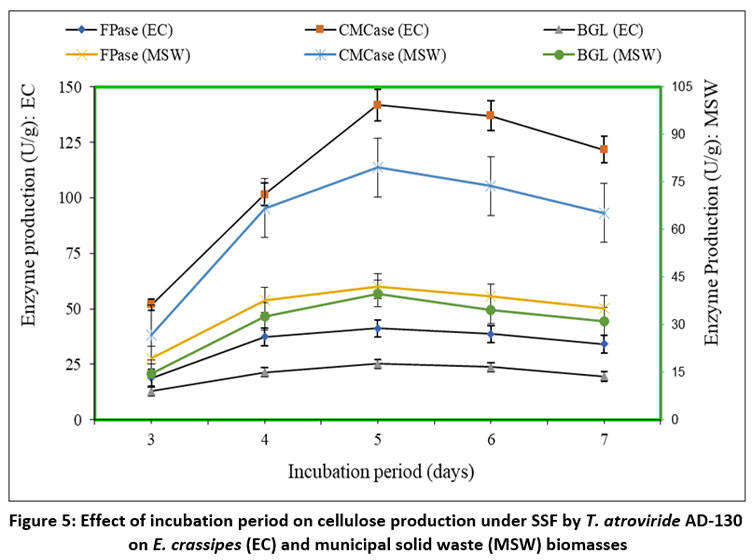 | Figure 5: Effect of incubation period on cellulose production under SSF by T. atroviride AD-130 on E. crassipes (EC) and municipal solid waste (MSW) biomasses
|
Fungi generally exhibit a broad pH tolerance, ranging from 4.0 to 8.0 (1998), and they have specific pH preferences for optimal growth and enzyme activity. To investigate the effect of initial pH on the cellulase activity using T. atroviride AD-130, the culture medium's pH was adjusted within the range of 4.0 to 8.0. Figure 6 illustrates that the highest yields of FPase (50.09U/g), CMCase (166.21U/g), and BGL (40.49U/g) were achieved at pH 7.0 during SSF of E. crassipes, using a substrate moisture ratio of 1:6 at 30°C, on the 5th day. At pH 4.0 and 8.0, the FPase yields were 40.06U/g and 42.19U/g, respectively. Likewise, CMCase activities at pH 4.0 and pH 8.0 were 147.57U/g and 79.04U/g, respectively, while the corresponding BGL yields were 28.09U/g and 31.03U/g.
Figure 6 also illustrates that the peak activities of FPase (52.57U/g), CMCase (103.21U/g), and BGL (68.12U/g) were attained at pH 7.0 during MSW solid state fermentation (SSF), employing a substrate moisture ratio of 1:2 at 30°C, on the 5th day. At pH levels of 4.0 and pH 8.0, the respective activities of FPase, CMCase, and BGL were 38.17U/g, 67.82U/g, and 37.68U/g, as well as 47.04U/g, 87.51U/g, and 57.07U/g, respectively.
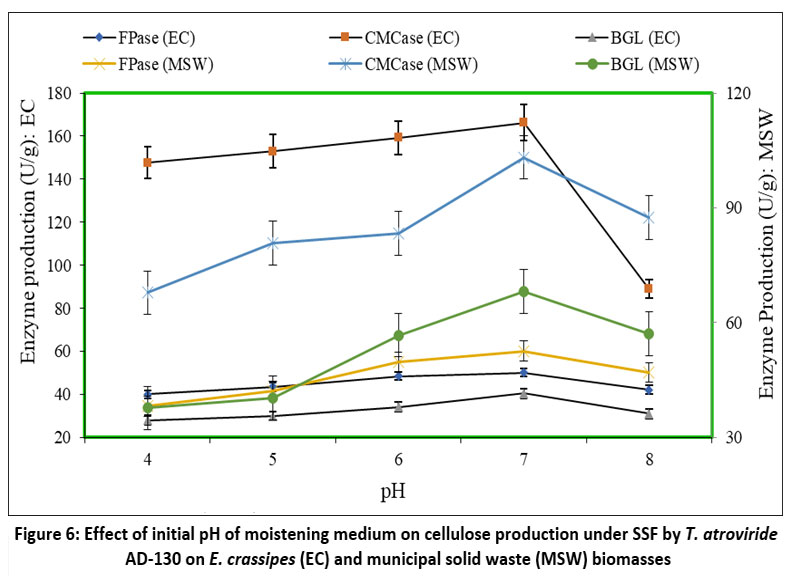 | Figure 6: Effect of initial pH of moistening medium on cellulose production under SSF by T. atroviride AD-130 on E. crassipes (EC) and municipal solid waste (MSW) biomasses
|
Impact of Nitrogen Sources on Cellulase Yield
Cellulase enzyme synthesis during SSF is influenced by the nature and concentration of nitrogen sources in the medium [41], as the fermentation process is significantly influenced by the pH of the solution, which can be altered by the nitrogen source employed [42]. Therefore, the current work aimed to evaluate the cellulase production of T. atroviride AD-130 during SSF, conducted at optimized 30°C and pH 7.0, under different combinations of organic and inorganic nitrogen sources (referred to as combinations I to VI in Table 2) with E. crassipes biomass and MSW residue serving as the substrates for a period of five days.
Table 2 illustrates the maximum cellulase production in term of FPase, CMCase and BGL activities by the fungus T. atroviride AD-130 as 52.57U/g, 103.03U/g, and 68.12U/g on MSW residue, and 50.09U/g, 166.21U/g, and 40.49U/g on E. crassipes biomass, respectively. These results were achieved during SSF when using a combination of 0.1% protease peptone, 0.14% ammonium sulfate, and 0.03% urea (referred to as combination I). Interestingly, altering the concentration of organic and inorganic nitrogen sources did not significantly contribute to the enhancement of cellulase production under SSF conditions. The lowest activities (U/g) of FPase, CMCase, and BGL, when using MSW residue and E. crassipes biomass as substrates, were observed in the presence of combination VI, with 0.2% protease peptone, 0.28% ammonium sulfate, and 0.03% urea, resulting in values of 40.82, 77.63, and 41.29, and 36.03, 118.71, and 27.09, respectively.
Table 2: Effect of various Nitrogen Sources on cellulase yields by T. atroviride AD-130 at 30°C and pH 7.0
Combination | Municipal solid wastes | E. crassipes | ||||
FPase Activity (U/g) | CMCase activity (U/g) | BGL activity (U/g) | FPase Activity (U/g) | CMCase activity (U/g) | BGL activity (U/g) | |
I | 52.57±2.47 | 103.21±5.03 | 68.12±3.41 | 50.09±2.25 | 166.21±6.28 | 40.49±1.28 |
II | 45.03±2.38 | 87.73±3.29 | 51.38±2.12 | 43.40±2.22 | 148.07±5.80 | 32.19±1.21 |
III | 41.63±1.67 | 86.98±3.70 | 49.31±1.82 | 41.82±1.94 | 143.15±3.78 | 30.91±1.27 |
IV | 49.42±2.56 | 97.83±5.09 | 63.05±2.27 | 47.15±2.87 | 157.47±6.21 | 38.03±1.37 |
V | 38.03±1.51 | 84.07±3.51 | 60.39±2.23 | 42.51±2.12 | 137.15±5.02 | 37.60±1.73 |
VI | 40.89±1.63 | 77.63±2.29 | 41.29±1.71 | 36.03±1.71 | 128.71±6.04 | 27.09±1.08 |
Combinations (I to VI)
I: 0.1% PP + 0.03% Urea + 0.14% AS
II: 0.05% PP + 0.03% Urea + 0.14% AS
III: 0.1% PP + 0.03% Urea+ 0.28% AS
IV: 0.1% PP + 0.03% Urea+ 0.05% YE + 0.21% AS
V: 0.05% PP + 0.03% Urea+ 0.1% YE + 0.21% AS
VI: 0.2% PP + 0.03% Urea+ 0.28% AS
AS: Ammonium Sulphate; PP: Protease Peptone; YE: Yeast Extract
Characteristics of Partially Purified Enzymes
Impact of pH on Enzyme Activity
The activity of enzymes is significantly influenced by the pH of the medium, as it affects the distribution of charges on both the substrate and enzyme molecules, thereby impacting catalysis through substrate binding to the enzyme's active site. Figure 7 illustrates how pH affects the performance of CMCase, FPase, and BGL. The optimal pH ranges for CMCase, FPase, and BGL were, 4.0 to 5.0, 5.0 to 7.0, and 4.5 to 5.5, with their respective optimum pH values being 4.5, 6.0, and 5.0. A marked reduction in relative enzyme activity was detected when the pH level ranged below and above the optimum level. The relative activities of FPase at pH 7.5 and 4.5 were 78.96% and 82.90%, respectively, compared to the optimum pH, while it was 73.84% at pH 3.5. CMCase exhibited relative activities of approximately 78% at pH 3.5 and 5.5, with a minimum relative activity of 18.79% at pH 7.5. The relative activities retained by B-glucosidase were 56.90% at pH 4.0, 63.59% at pH 6.0, and a minimum of 8.16% at pH 7.5.
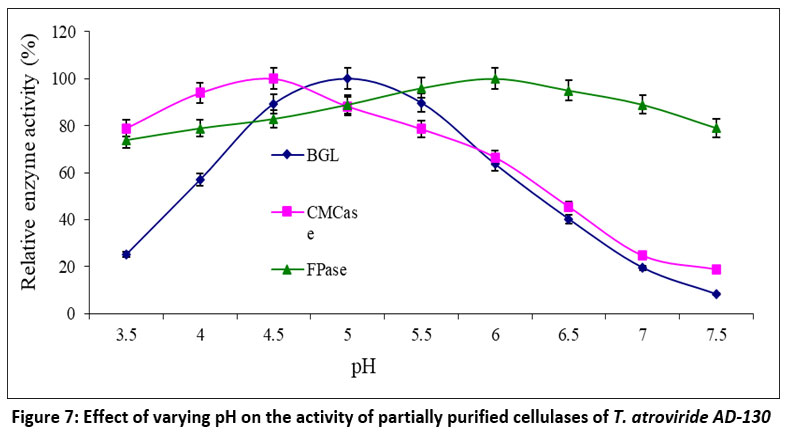 | Figure 7: Effect of varying pH on the activity of partially purified cellulases of T. atroviride AD-130.
|
Impact of Temperature on Enzyme Activity
Cellulases typically exhibit optimal activity at temperatures between 40°C to 60°C [52]. To determine the optimum temperature for FPase, CMCase, and BGL, enzyme samples were incubated with their respective substrates at temperatures ranging from 30°C - 80°C. The temperature dependence of FPase, CMCase, and BGL activities is presented in Figure 8. In this study, the optimum temperatures for FPase, CMCase, and BGL activities was investigated as 60°C, 50°C, and 70°C, respectively. The respective optimal temperature ranges for FPase, CMCase, and BGL were found to be 40 to 70°C, 40 to 60°C, and 60 to 70°C. Significantly, at 80°C, there was a noticeable reduction in FPase and BGL activity, whereas CMCase activity decreased at 70°C, resulting in residual activities of 62.17%, 41.88%, and 48.68%, respectively, at these temperatures.
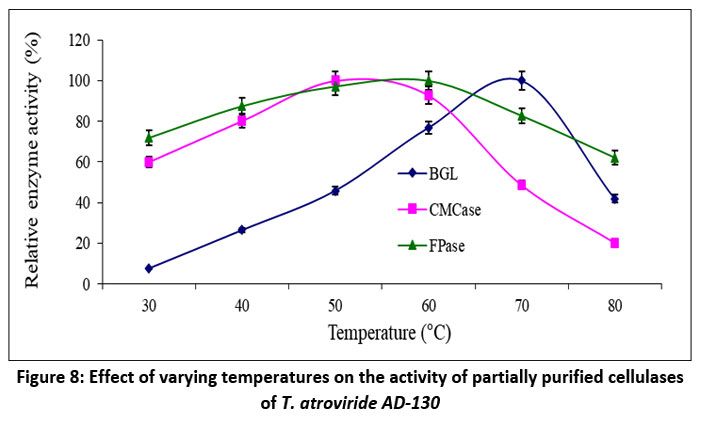 | Figure 8: Effect of varying temperatures on the activity of partially purified cellulases of T. atroviride AD-130
|
Detection of CMCase and B-glucosidase Activity by Zymogram Staining
In the current study, the zymograms of CMCase and BGL were prepared by incorporating 0.2% carboxymethylcellulose (CMC) and 0.5% p-nitrophenol-D-glucopyranoside into 10% native PAGE respectively as shown in Figure 9. Zymogram analysis of partially purified cellulases from T. atroviride AD-130 exhibited six distinct bands (C1, C2, C3, C4, C5, and C6) corresponding to different isoforms of CMCase, along with a single active band (G1) for B-glucosidase.
 | Figure 9: Zymogram staining on native PAGE of partially purified CMCase and B-glucosidase
|
Discussion
Different studies reported varying optimal substrate-to-water ratios and moisture levels for maximizing enzyme production. Researchers 12 found a 1:1.5 ratio suitable for highest CMCase and FPase production by Aspergillus niger, while a 1:1.75 ratio yielded the highest B-glucosidase yield. A study demonstrated that Trichoderma asperellum achieved maximum FPase production (26.03U/g) at 60-80% moisture content during SSF of oil palm frond leaves 32. Another study reported the maximum cellulase production at a 70% moisture level using A. niger 17. Similarly, the highest FPase yield using A. niger and T. reesei at 70% moisture level was obtained33. Talaromyces stipitatus exhibited highest FPase (4.51FPU/gds), endoglucanase (53.3IU/gds), and BGL (62.6IU/gds) activities with Parthenium hysterophorous as substrate under SSF conditions (pH 6.0, 30°C) with 80% moisture level34. Thus, multiple studies have emphasized the impact of moisture levels on cellulase enzymes across various fungal strains and substrates. In this study, deviations from the optimal moisture content, whether higher or lower, negatively affected enzyme production because the increased moisture content might have led to decreased porosity, changes in substrate structure, reduced gas volume, and hindered inter-particle spaces35 while the lower moisture content could have reduced nutrient solubility, impeded swelling, and increase water tension within the solid substrate.
The cellulase production in the present study exhibited a pattern where the minimum production was observed on the 3rd day, followed by a peak on the 5th day, and a subsequent decline. This decline in enzyme yields could be attributed to the release of proteases and a decrease in the pH of the medium, which, as suggested by the previous study36, might have been influenced by various factors, including the ratios of amorphous to crystalline cellulose. Herewith, it is also noteworthy that some of the studies testified the highest cellulases production by A. niger after 120 hours of incubation17, whereas some work contradicts the results and achieved significant cellulases production within a three-day fermentation period using Aspergillus and Trichoderma spp37,38. Likewise, researchers reported maximum FPase and B-glucosidase (BGL) production by A. niger after 96 hours of fermentation39 while some authors observed the highest FPase and CMCase production by T. harzianum on the 4th day of fermentation 40. These contrasting results highlight the variability in cellulases production kinetics among different fungal strains and underscore the importance of optimizing fermentation conditions for each specific strain to achieve maximum enzyme yields.
The obtained results align with previous works, which also observed that the highest yields of CMCase, FPase, and BGL were achieved at pH 7.0 12,13. Contrarily, a study reported that A. niger exhibited the highest cellulase production at an initial pH of 5.017. Similarly, rediscovered that A. niger displayed the highest production of FPase and B-glucosidase at an initial pH of 4.039. At the same time, maximum production of FPase and CMCase at an initial pH of 6.0 was observed by T. harzianum 40.
The study demonstrated that the maximum yield using A. flavus was achieved via incorporating 2 g/L sodium nitrate (NaNO3T) and 2 g/L protease peptone as nitrogen sources in the fermentation medium43. In contrast, highest cellulase yield by A. terreus when fermentation medium contained 1% (w/v) yeast extract44. Additionally, studies have found that inorganic nitrogen sources to be optimal for cellulase production. For instance, researcher reported that, T. harzianum exhibited maximum enzyme production of CMCase (26.22IU/gds) and total cellulase (7.42FPU/gds) in the presence of 0.02M ammonium ferrous sulfate40. Similarly, a study found that incorporating 0.2% (w/w) urea in the fermentation medium resulted in maximum cellulase production by A. niger39. Study reveals that, addition of urea and ammonium sulfate significantly increased cellulase production by P. funiculosum 41. In optimizing the medium components for cellulase synthesis by Melanocarpus sp. under SSF, study found that the addition of urea to rice straw, used as the carbon source, was beneficial45. However, one study observed no significant increase in cellulase activity in A. niger when using any of the investigated organic or inorganic nitrogen sources17.
Previous studies have also reported optimal pH values for CMCase and FPase produced by T. atroviride to be 4.0 and 5.0, respectively46. In another study pH 5.5 was found to be the most favorable for CMCase activity in Trichoderma sp.47, while pH 5.0 was identified as the optimal condition for FPase activity in another strain of T. atroviride 48. Although endoglucanase and B-glucosidase produced by A. terreus exhibited higher activity at pH 2.0 and pH 3.0, respectively44, but pH 7.0 found to be the most favorable for CMCase activity49. A. niger's CMCase exhibits its most effective performance in the pH of 6.0 to 7.050. The optimal pH values for CMCase, B-glucosidase, and FPase produced by Aspergillus fumigatus were determined to be pH 3.0, 4.0, and 5.0, respectively51. In another work, pH 4.0 reported as the most favourable for cellulase activity in A. niger17 . However, the optimal pH values for CMCase and FPase produced by T. harzianum to be 5.5 and 5.0, respectively40.
CMCase and FPase produced by T. atroviride displayed optimal activity at 60°C and 50°C, respectively (47). Most endoglucanases were found to be optimally active at temperatures close to 50°C 50. However, maximum FPase activity was reported from T. atroviride at 50°C 46, while the optimal temperature for FPase activity identified as 60°C in Trichoderma sp. A-00153. CMCase and BGL were also found to be optimally active at 70°C, while FPase exhibited optimal activity at 60°C54. Both endoglucanase (CMCase) and B-glucosidase from A. terreus displayed maximum activity at 70°C44. Cellulase produced by A. niger demonstrated peak activity at 65°C 17. In line with these findings, a study was reported the optimal temperatures for CMCase and FPase synthesized by T. harzianum to be 60°C and 55°C, respectively 40.
Similarly, zymogram analysis of A. niger enzyme extracts revealed six active bands for CMCase17. In contrast, researchers identified two active bands for CMCase in T. atroviride zymogram staining, indicating the presence of two isoforms46, while team of authors observed three active bands for CMCase and one band for B-glucosidase in cellulases produced by T. viride54. Zymogram analysis of A. fumigates cellulases indicated the presence of three CMCase isoforms and two B-glucosidase isoforms51. Additionally, native PAGE for characterization of partially purified enzymes from A. flavus observed a single activity band for CMCase43.
Conclusion
To optimize cellulase production during SSF, the newly isolated T. atroviride AD-130 was cultivated using untreated and dried lignocellulosic biomasses, namely E. crassipes and municipal solid waste, at 30°C. The highest cellulase production was attained on the 5th day, with a substrate-moisture ratio of 1:6 for E. crassipes and 1:2 for MSW, with pH attuned to 7.0. Optimized medium contained 0.1% protease peptone, 0.14% ammonium sulfate, and 0.03% urea, with a pH range of 6.0-6.5 for FPase, 4.5-5.0 for CMCase, and 5.0-5.5 for B-glucosidase activity. The optimum temperatures for FPase, CMCase, and BGL were determined as 60°C, 50°C, and 70°C, respectively. Zymogram of partially purified cellulases from T. atroviride AD-130 exhibited six isoforms (C1, C2, C3, C4, C5, and C6) of CMCase, along with a single active band (G1) for B-glucosidase. Despite the limited B-glucosidase production in numerous Trichoderma spp., T. atroviride AD-130 demonstrated a well-balanced proportion of both FPase and BGL. This balanced enzyme profile will be quite conducive for efficient cellulose hydrolysis into glucose by overcoming cellobiose inhibition.
Acknowledgement
All the participants of the cited paper acknowledge the Major Research Project funded by the University Grants Commission; New Delhi (India).
Funding Sources
University Grants Commission; New Delhi (India) vide F. No. 35-45/2008 (SR) dated 01/05/2009
Conflict of Interest
The author(s )do not have any conflict of interest.
Data Availability Statement
This statement does not apply to this article
Ethics Statement
This research did not involve human participants, animal subjects, or any material that requires ethical approval
Informed Consent Statement
This study did not involve human participants, and therefore, informed consent was not required.”
Author Contributions
Anil Kumar: Ph.D. student and Co-investigator, of Research Project.
Dr. Meenakshi Suhag: helped draft, edit, and corresponding author manuscripts.
Dr. Joginder Singh: Principal Investigator of Research Project, Conceptualization, Methodology, Designing and Editing
Dr. Ritu Nandal: Assist in Experimental Work
Dr. Naveeta Dhaka: Assist in Experimental Work
Dr. Rajesh Dhankhar: Supervisor of Co-investigator
References
- Larnaudie V, Ferrari M.D, Lareo C. Switchgrass as an alternative biomass for ethanol production in a biorefinery: Perspectives on technology, economics and environmental sustainability. Renewable & Sustainable Energy Reviews 2022; 158:112115.
CrossRef
CrossRef- Santos G.B, de Sousa Francisco Filho Á, da Silva Rodrigues J.R, de Souza R.R. Cellulase production by Aspergillus niger using urban lignocellulosic waste as substrate: Evaluation of different cultivation strategies, Journal of Environmental Management 2022; (305) 114431.
CrossRef - Abduh M.Y, Ramadhan C.R, Fadhlilah A.P, Abdul S.D, Burhan K.H. Solid-state fermentation of groundnut (Arachis hypogaea) shell using Trichoderma sp., tape yeast, and tempeh yeast to produce cellulase, J. Appl. Biol. Biotechnol 2022; 10(4) 153-160.
CrossRef - Chysirichote T, Phaiboonsilpa N, Laosiripojana N. High production of cellulase and xylanase in solid-state fermentation by Trichoderma reesei using spent copra and wheat bran in rotary bioreactor, Industrial & Engineering Chemistry Research 2023; 62(7) 3087-3097.
CrossRef - Santos F.A, de Carvalho-Gonçalves L.C, de Carvalho Cardoso-Simões A.L, de Melo Santos S.F. Evaluation of the production of cellulases by Penicillium sp. FSDE15 using corncob and wheat bran as substrates, Bioresource Technology Reports 2021; (14)100648.
CrossRef - Verma N, Kumar V. Impact of process parameters and plant polysaccharide hydrolysates in cellulase production by Trichoderma reesei and Neurospora crassa under wheat bran based solid state fermentation, Biotechnology reports 2020; (25) e00416.
CrossRef - Dinil A, Jacob A.Valorization of Agro-industrial Discards in Fermentation for the Production of Cellulase Enzyme, Journal of Pure & Applied Microbiology 2022;16(1).
CrossRef - Sankar M, Mathew R.M, Puthiyamadam A, Sreeja-Raju A, Christopher M, Gokhale D.V, Sukumaran R.K. Comparison of the solid-state and submerged fermentation derived secretomes of hyper-cellulolytic Penicillium janthinellum NCIM 1366 reveals the changes responsible for differences in hydrolytic performance, Bioresource Technology 2023; (371)128602.
CrossRef - Teigiserova D.A, Bourgine J, Thomsen M. Closing the loop of cereal waste and residues with sustainable technologies: An overview of enzyme production via fungal solid-state fermentation, Sustainable Production and Consumption 2021; (27) 845-857.
CrossRef - Behera S.S, Ray R.C. Solid state fermentation for production of microbial cellulases: recent advances and improvement strategies, International journal of biological macromolecules 2016; (86) 656-669.
CrossRef - Lodha A, Pawar S, Rathod V. Optimised cellulase production from fungal co-culture of Trichoderma reesei NCIM 1186 and Penicillium citrinum NCIM 768 under solid state fermentation, Journal of Environmental Chemical Engineering 2020; 8(5) 103958.
CrossRef - Alam M.Z, Muhammad N, Mahmat M.E. Production of cellulase enzyme from oil palm biomass as substrate by solid state bioconversion, Am. J. Appl. Sci 2005; (2) 569-572.
CrossRef - Karmakar M, Ray R.R. Extra cellular endoglucanase production by Rhizopus oryzae in solid and liquid state fermentation of agro wastes", Asian J Biotechnol 2010; 2 (1) 27-36.
CrossRef - Juwaied A.A, Adnan S, Al-Amiery A.A. Production cellulase by different co-culture of Aspergillus niger and Trichoderma viride from waste paper, Journal of Yeast and Fungal Research 2010; 1(6) 108-111.
- Bansal N, Tewari R, Soni R, Soni S.K. Production of cellulases from Aspergillus niger NS-2 in solid state fermentation on agricultural and kitchen waste residues, Waste management 2012; 32(7) 1341-1346.
CrossRef - Ncube T, Howard R.L, Abotsi E.K, van Rensburg E.L, Ncube I. Jatropha curcas seed cake as substrate for production of xylanase and cellulase by Aspergillus niger FGSCA733 in solid-state fermentation, Industrial Crops and Products 2012; 37 (1) 118-123.
CrossRef - Molaverdi M, Karimi K, Khanahmadi M, Goshadrou A. Enhanced sweet sorghum stalk to ethanol by fungus Mucor indicus using solid state fermentation followed by simultaneous saccharification and fermentation, Industrial crops and products 2013; (49) 580-585.
CrossRef - Maza M, Pajot H.F, Amoroso M.J, Yasem M.G. Post-harvest sugarcane residue degradation by autochthonous fungi, International Biodeterioration & Biodegradation 2014; (87) 18-25.
CrossRef - Narra M, Dixit G, Divecha J, Kumar K, Madamwar D, Shah A.R. Production, purification and characterization of a novel GH 12 family endoglucanase from Aspergillus terreus and its application in enzymatic degradation of delignified rice straw, In Biodeterior Biodegrad 2014; 150-161.
CrossRef - Marín M, Sánchez A, Artola A. Production and recovery of cellulases through solid-state fermentation of selected lignocellulosic wastes, Journal of cleaner production 2019; (209) 937-946.
CrossRef - Li J, Tang X, Chen S, Zhao J, Shao T. Ensiling pretreatment with two novel microbial consortia enhances bioethanol production in sterile rice straw, Bioresource technology 2021; (339) 125507.
CrossRef - Areeshi MY. Microbial cellulase production using fruit wastes and its applications in biofuels production, International Journal of Food Microbiology 2022; (378) 109814.
CrossRef - Li C, Yang Z, Zhang R.H, Zhang D, Chen S, Ma L. Effect of pH on cellulase production and morphology of Trichoderma reesei and the application in cellulosic material hydrolysis, Journal of Biotechnology 2013; 168(4) 470-477.
CrossRef - Das S.P, Gupta A, Das D, Goyal A. Enhanced bioethanol production from water hyacinth (Eichhornia crassipes) by statistical optimization of fermentation process parameters using Taguchi orthogonal array design, International biodeterioration & biodegradation 2016; (109) 174-184.
CrossRef - Abdel-Fattah A.F, Abdel-Naby M.A. Pretreatment and enzymic saccharification of water hyacinth cellulose, Carbohydrate polymers 2012; 87(3), 2109-2113.
CrossRef - Clark H.E, Geldrich E.F, Kabler P.W, Huff C.B. Applied microbiology” International Book Company, New York, 53.
- Mandels M, Sternberg D.J. Recent advances in cellulose technology, J Ferment Technol 1976; (54) 267-286.
- Sternberg D.A, Mandels G. Induction of cellulolytic enzymes in Trichoderma reesei by sophorose, Journal of Bacteriology 1979; 139(3) 761-769.
CrossRef - Ghose T.K. Measurement of cellulase activities, Pure Appl Chem 1987; (59) 257–268.
CrossRef - Ghose T.K, Bisaria V.S. Measurement of Hemicellulase Activities, Part I- Xylanases, Pure AppI Chem 1987; (59) 1739-1751.
CrossRef - Ezeilo U.R, Lee C.T, Huyop F, Zakaria I.I, Wahab R.A. Raw oil palm frond leaves as cost-effective substrate for cellulase and xylanase productions by Trichoderma asperellum UC1 under solid-state fermentation, Journal of Environmental Management 2019; (243) 206-217.
CrossRef - Lee C.K, Darah I, Ibrahim C.O. Production and optimization of cellulase enzyme using Aspergillus niger USM AI 1 and comparison with Trichoderma reesei via solid state fermentation system, Biotechnology Research International 2011; (1), 658493.
CrossRef - Bharti A.K, Kumar A, Kumar A, Dutt D. Exploitation of Parthenium hysterophorous biomass as low-cost substrate for cellulase and xylanase production under solid-state fermentation using Talaromyces stipitatus MTCC 12687, Journal of radiation research and applied sciences 2018; 11(4) 271-280.
CrossRef - Anto H, Trivedi U.B, Patel K.C. Glucoamylase production by solid-state fermentation using rice flake manufacturing waste products as substrate, Bioresource Technology 2006; 97(10) 1161-1166.
CrossRef - Ögel Z.B, Yarangümeli K, Dündar H, Ifrij I. Submerged cultivation of Scytalidium thermophilum on complex lignocellulosic biomass for endoglucanase production, Enzyme and microbial technology 2001; 28(7-8) 689-695.
CrossRef - Chandra M.S, Viswanath B, Reddy B.R. Cellulolytic enzymes on lignocellulosic substrates in solid state fermentation by Aspergillus niger, Indian Journal of Microbiology 2007; (47) 323-328.
CrossRef - Chandra M.S, Reddy B.R. Exoglucanase production by Aspergillus niger grown on wheat bran, Ann Microbiol 2013; 63(3) 871-7.
CrossRef - Oberoi H.S, Rawat R, Chadha B.S. Response surface optimization for enhanced production of cellulases with improved functional characteristics by newly isolated Aspergillus niger HN-2. Antonie van Leeuwenhoek 2014; (105) 119-134.
CrossRef - Pathak P, Bhardwaj N.K, Singh A.K. Production of crude cellulase and xylanase from Trichoderma harzianum PPDDN10 NFCCI-2925 and its application in photocopier waste paper recycling, Applied biochemistry and biotechnology 2014; (172) 3776-3797.
CrossRef - Maeda R.N, da Silva M.M, Santa Anna L.M, Pereira N. Nitrogen source optimization for cellulase production by Penicillium funiculosum, using a sequential experimental design methodology and the desirability function, Applied Biochemistry and Biotechnology 2010; (161) 411-422.
CrossRef - Kachlishvili E, Penninckx M.J, Tsiklauri N, Elisashvili V. Effect of nitrogen source on lignocellulolytic enzyme production by white-rot basidiomycetes under solid-state cultivation, World Journal of Microbiology and Biotechnology 2006; (22) 391-397.
CrossRef - Sajith S, Sreedevi S, Priji P, Unni K.N, Benjamin S. Production and partial purification of cellulase from a novel fungus, Aspergillus flavus BS1, Annals of Microbiology 2014; (64) 763-771.
CrossRef - Gao J, Weng H, Zhu D, Yuan M, Guan F, Xi Y. Production and characterization of cellulolytic enzymes from the thermoacidophilic fungal Aspergillus terreus M11 under solid-state cultivation of corn stover, Bioresource technology 2008; 99(16) 7623-7629.
CrossRef - Jatinder K.A, Chadha B.S, Saini H.S. Optimization of medium components for production of cellulases by Melanocarpus sp. MTCC 3922 under solid-state fermentation, World Journal of Microbiology and Biotechnology 2006; (22) 15-22.
CrossRef - Grigorevski-Lima A.L, de Oliveira M.M, do Nascimento R.P, Bon E.P, Coelho R.R. Production and partial characterization of cellulases and xylanases from Trichoderma atroviride 676 using lignocellulosic residual biomass, Applied biochemistry and biotechnology 2013; (169) 1373-1385.
CrossRef - Adsul M.G, Bastawde K.B, Varma A.J, Gokhale D.V. Strain improvement of Penicillium janthinellum NCIM 1171 for increased cellulase production. Bioresource Technology 2007; 98(7) 1467-1473.
CrossRef - Kovács K, Szakacs G, Zacchi G. Comparative enzymatic hydrolysis of pretreated spruce by supernatants, whole fermentation broths and washed mycelia of Trichoderma reesei and Trichoderma atroviride, Bioresource Technology 2009; 100(3) 1350-1357.
CrossRef - Gomathi D, Muthulakshmi C, Kumar D.G, Ravikumar G, Kalaiselvi M, Uma C. Submerged fermentation of wheat bran by Aspergillus flavus for production and characterization of carboxy methyl cellulase, Asian Pacific Journal of Tropical Biomedicine 2012; 2(1) S67-S73.
CrossRef - Akiba S, Kimura Y, Yamamoto K, Kumagai H. Purification and characterization of a protease-resistant cellulase from Aspergillus niger, Journal of fermentation and bioengineering 1995; 79(2) 125-130.
CrossRef - Ang S.K, Shaza E.M, Adibah Y, Suraini A.A, Madihah M.S. Production of cellulases and xylanase by Aspergillus fumigatus SK1 using untreated oil palm trunk through solid state fermentation, Process Biochemistry 2013; 48(9) 1293-1302.
CrossRef - Dutta T, Sahoo R, Sengupta R, Ray S.S, Bhattacharjee A, Ghosh S. Novel cellulases from an extremophilic filamentous fungi Penicillium citrinum: production and characterization, Journal of Industrial Microbiology and Biotechnology 2008; 35(4) 275-282.
CrossRef - Gashe BA. Cellulase production and activity by Trichoderma sp. A?001, Journal of Applied Bacteriology 1992; 73(1) 79-82.
CrossRef - Zhou J, Wang Y.H, Chu J, Zhuang Y.P, Zhang S.L, Yin P. Identification and purification of the main components of cellulases from a mutant strain of Trichoderma viride T 100-14", Bioresource Technology 2008; 99(15) 6826-6833.
CrossRef







